

Acts of appetite: neural circuits governing the appetitive, consummatory, and terminating phases of feeding
- PMID: 35879462
- PMCID: PMC10852214
- DOI: 10.1038/s42255-022-00611-y
Acts of appetite: neural circuits governing the appetitive, consummatory, and terminating phases of feeding
Abstract
The overconsumption of highly caloric and palatable foods has caused a surge in obesity rates in the past half century, thereby posing a healthcare challenge due to the array of comorbidities linked to heightened body fat accrual. Developing treatments to manage body weight requires a grasp of the neurobiological basis of appetite. In this Review, we discuss advances in neuroscience that have identified brain regions and neural circuits that coordinate distinct phases of eating: food procurement, food consumption, and meal termination. While pioneering work identified several hypothalamic nuclei to be involved in feeding, more recent studies have explored how neuronal populations beyond the hypothalamus, such as the mesolimbic pathway and nodes in the hindbrain, interconnect to modulate appetite. We also examine how long-term exposure to a calorically dense diet rewires feeding circuits and alters the response of motivational systems to food. Understanding how the nervous system regulates eating behaviour will bolster the development of medical strategies that will help individuals to maintain a healthy body weight.
© 2022. This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply.
Figures
References
-
-
Sternson SM & Eiselt AK Three pillars for the neural control of appetite. Annu Rev. Physiol 79, 401–423 (2017).
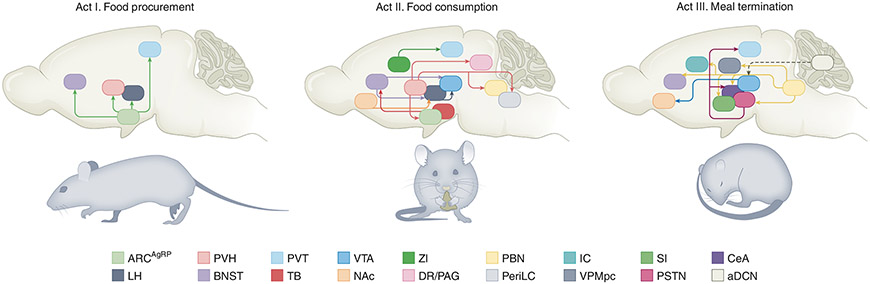
This review served as inspiration for dividing up the discrete phases of feeding into three acts.
-
-
- Aravich PF & Sclafani A Paraventricular hypothalamic lesions and medial hypothalamic knife cuts produce similar hyperphagia syndromes. Behav. Neurosci 97, 970–983 (1983). - PubMed
-
- Bergen HT, Mizuno TM, Taylor J & Mobbs CV Hyperphagia and weight gain after gold-thioglucose: relation to hypothalamic neuropeptide Y and proopiomelanocortin. Endocrinology 139, 4483–4488 (1998). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
