

Pseudorabies Virus Tegument Protein UL13 Suppresses RLR-Mediated Antiviral Innate Immunity through Regulating Receptor Transcription
- PMID: 35891444
- PMCID: PMC9317333
- DOI: 10.3390/v14071465
Pseudorabies Virus Tegument Protein UL13 Suppresses RLR-Mediated Antiviral Innate Immunity through Regulating Receptor Transcription
Abstract
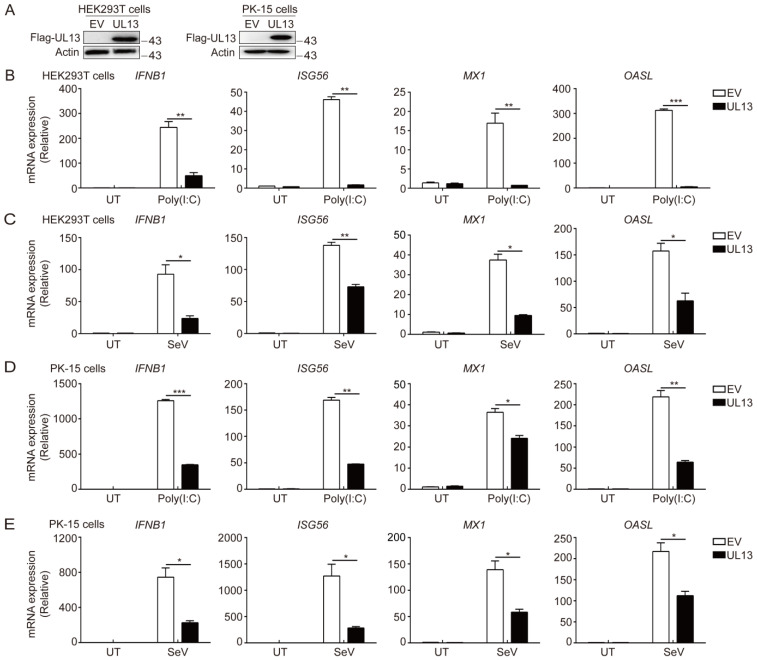
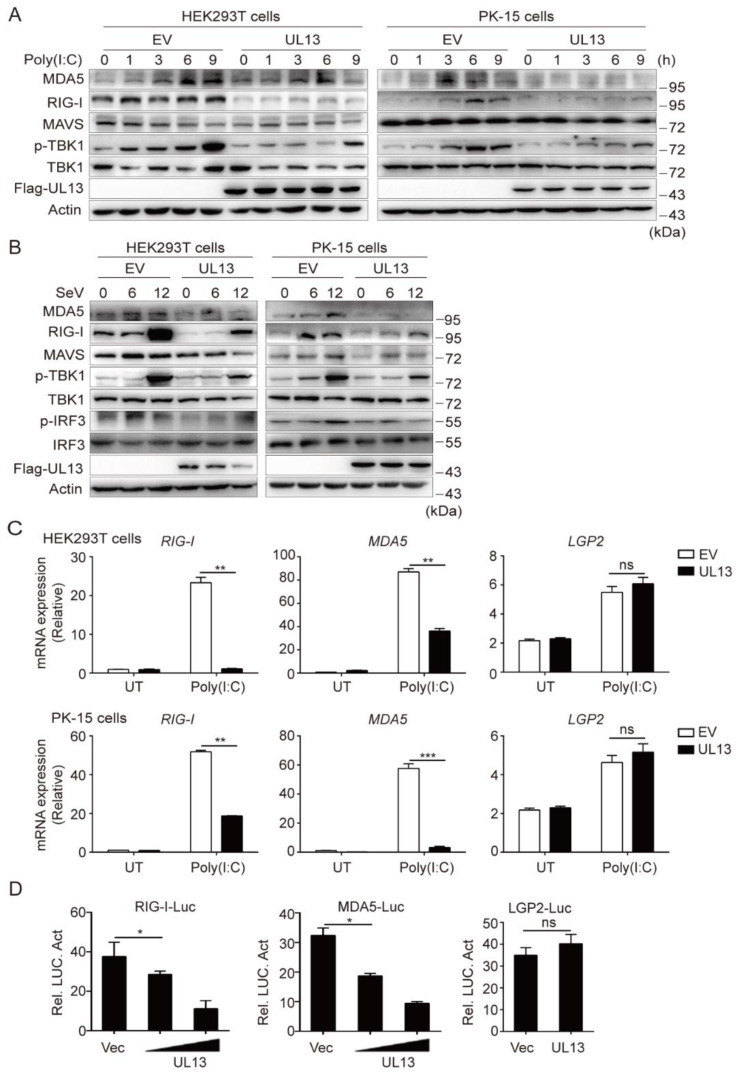
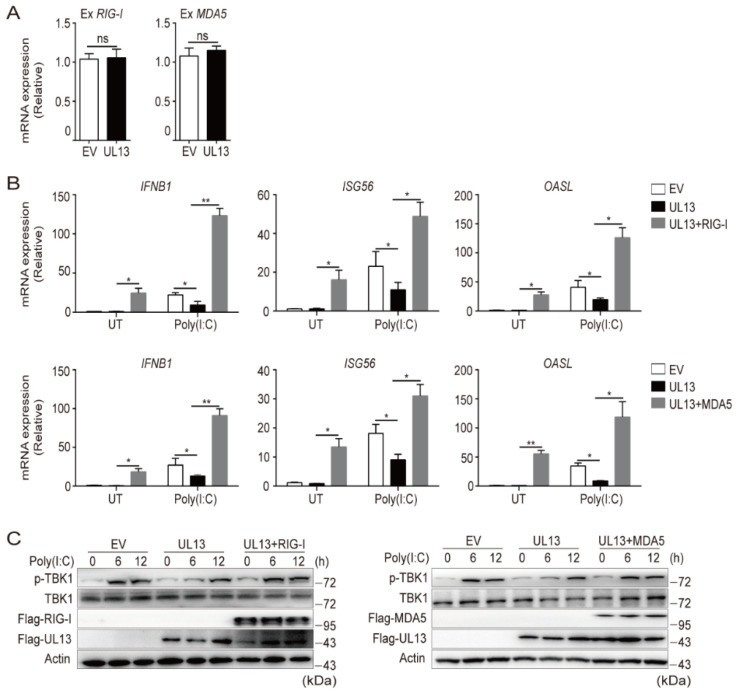
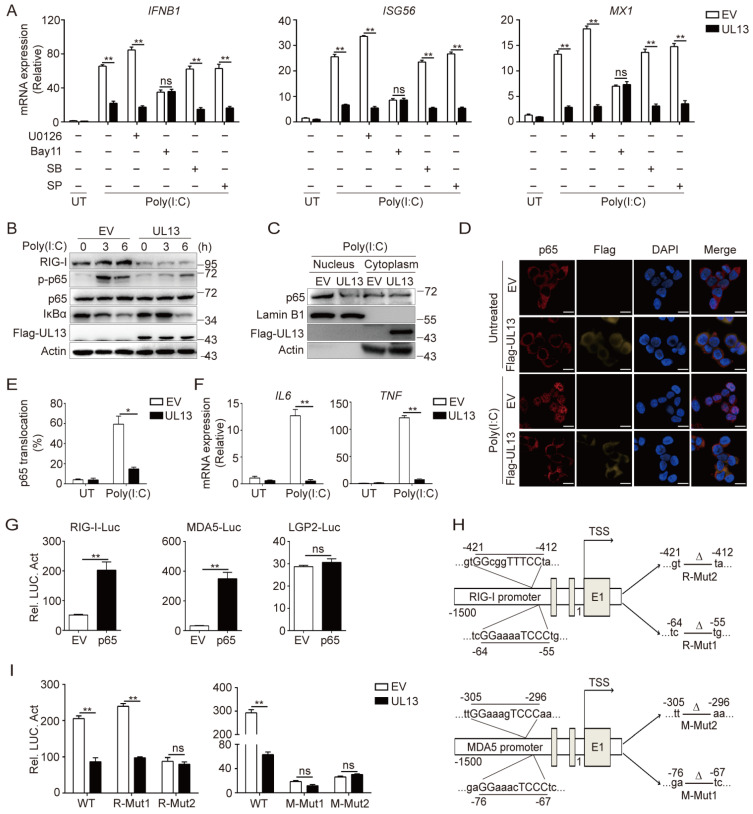
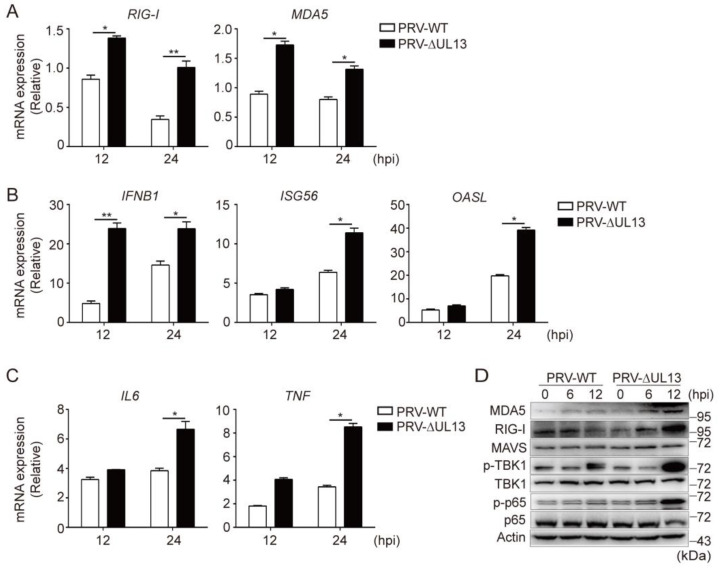
Pseudorabies virus (PRV) has evolved various strategies to escape host antiviral immune responses. However, it remains unclear whether and how PRV-encoded proteins modulate the RIG-I-like receptor (RLR)-mediated signals for immune evasion. Here, we show that the PRV tegument protein UL13 functions as an antagonist of RLR-mediated antiviral responses via suppression of the transcription of RIG-I and MDA5, but not LGP2. UL13 overexpression significantly inhibits both the mRNA and protein levels of RIG-I and MDA5, along with RIG-I- or MDA5-mediated antiviral immune responses, whereas overexpression of RIG-I or MDA5 counteracts such UL13-induced suppression. Mechanistically, UL13 suppresses the expression of RIG-I and MDA5 by inhibiting activation of the transcription factor NF-κB. Consequently, overexpression of p65 promotes the activation of RIG-I and MDA5 promoters. Moreover, deletion of the p65-binding sites in the promoters of RIG-I or MDA5 abolishes the suppression role of UL13. As a result, mutant PRV lacking UL13 elicits stronger host antiviral immune responses than PRV-WT. Hence, our results provide a novel functional role of UL13-induced suppression of host antiviral immunity through modulating receptors' transcription.
Keywords: MDA5; NF-κB; RIG-I; pseudorabies virus (PRV); tegument protein UL13.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
