

EGFR signaling activates intestinal stem cells by promoting mitochondrial biogenesis and β-oxidation
- PMID: 35896119
- PMCID: PMC10117080
- DOI: 10.1016/j.cub.2022.07.003
EGFR signaling activates intestinal stem cells by promoting mitochondrial biogenesis and β-oxidation
Abstract
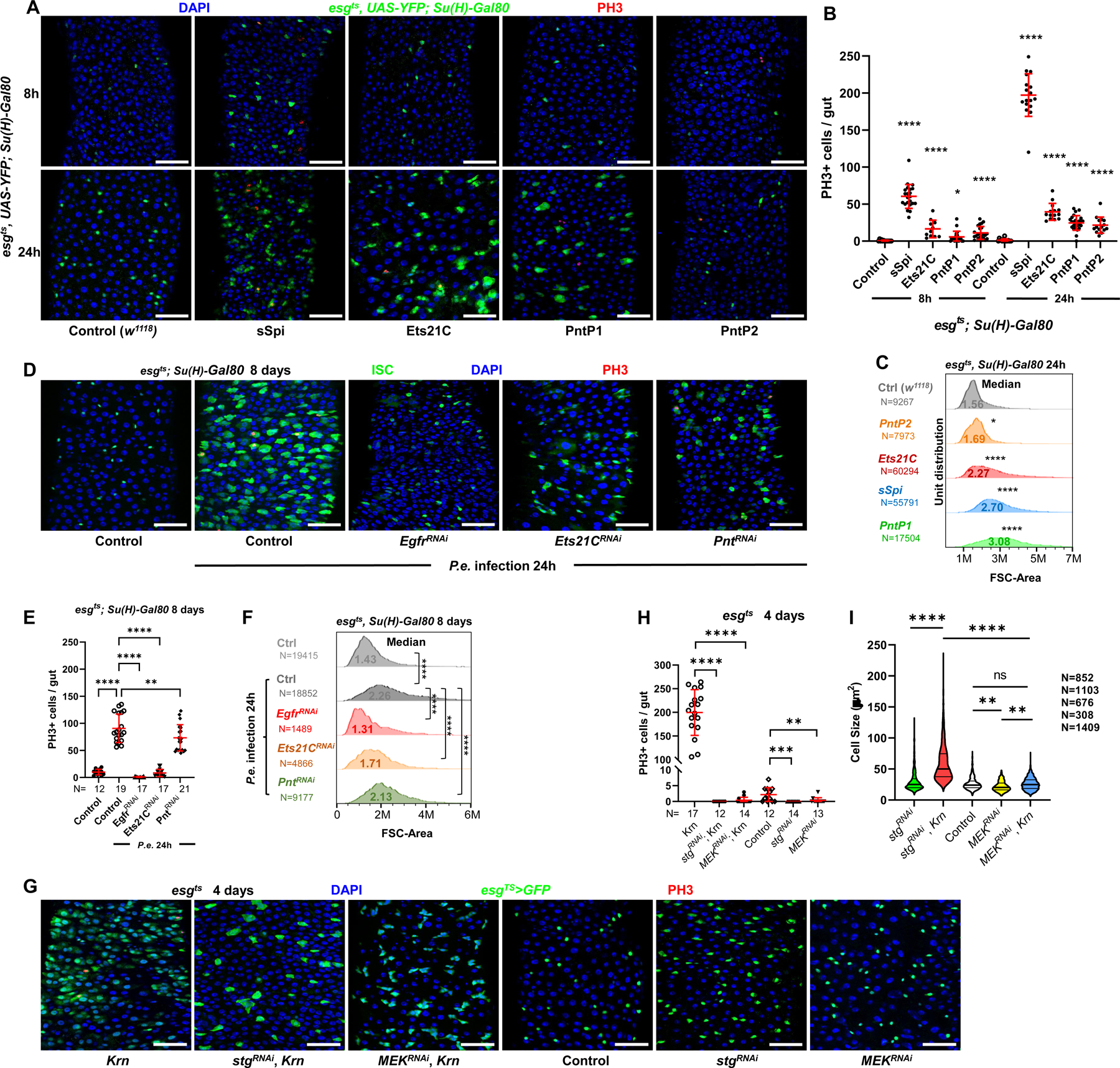
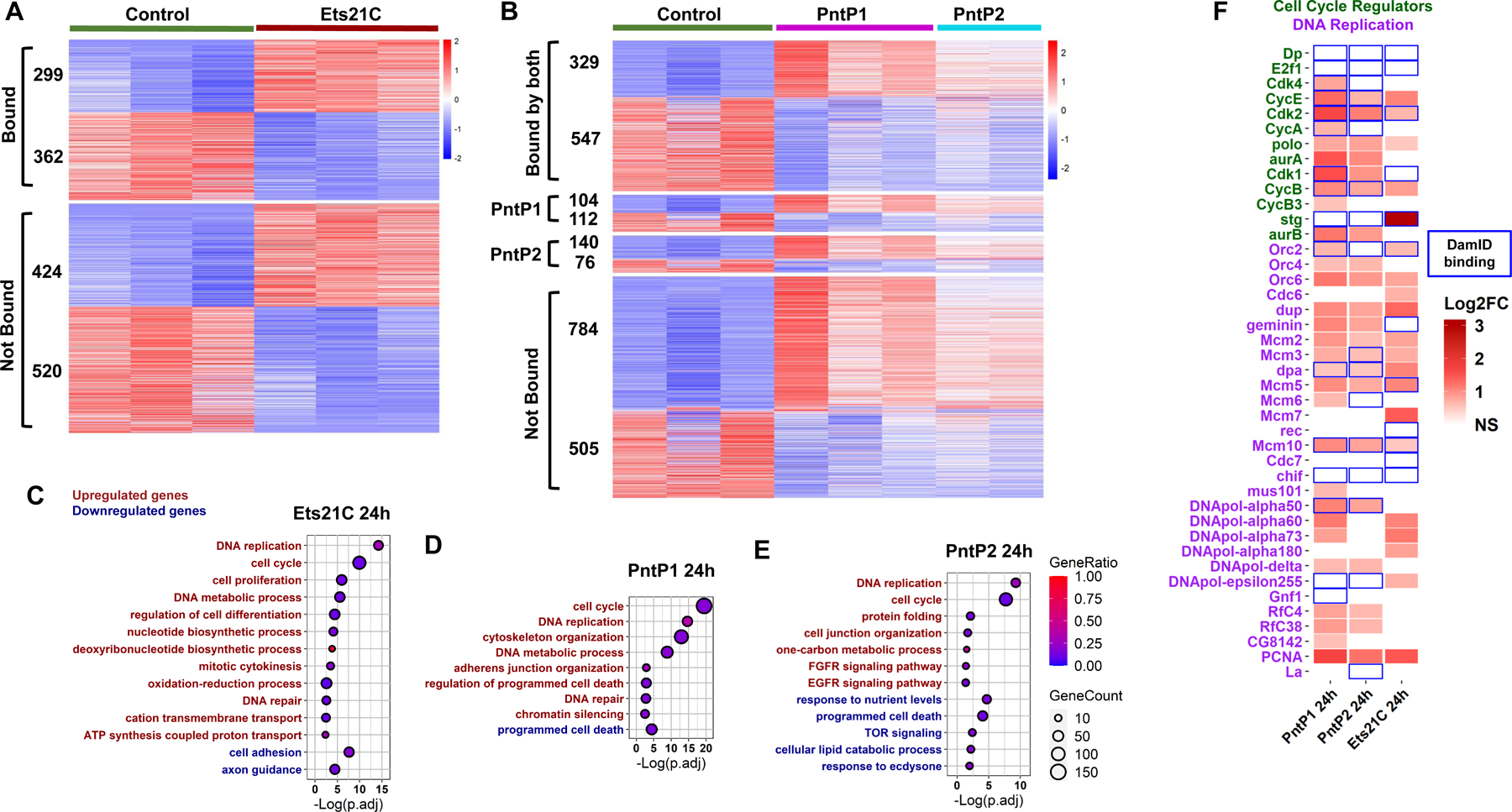
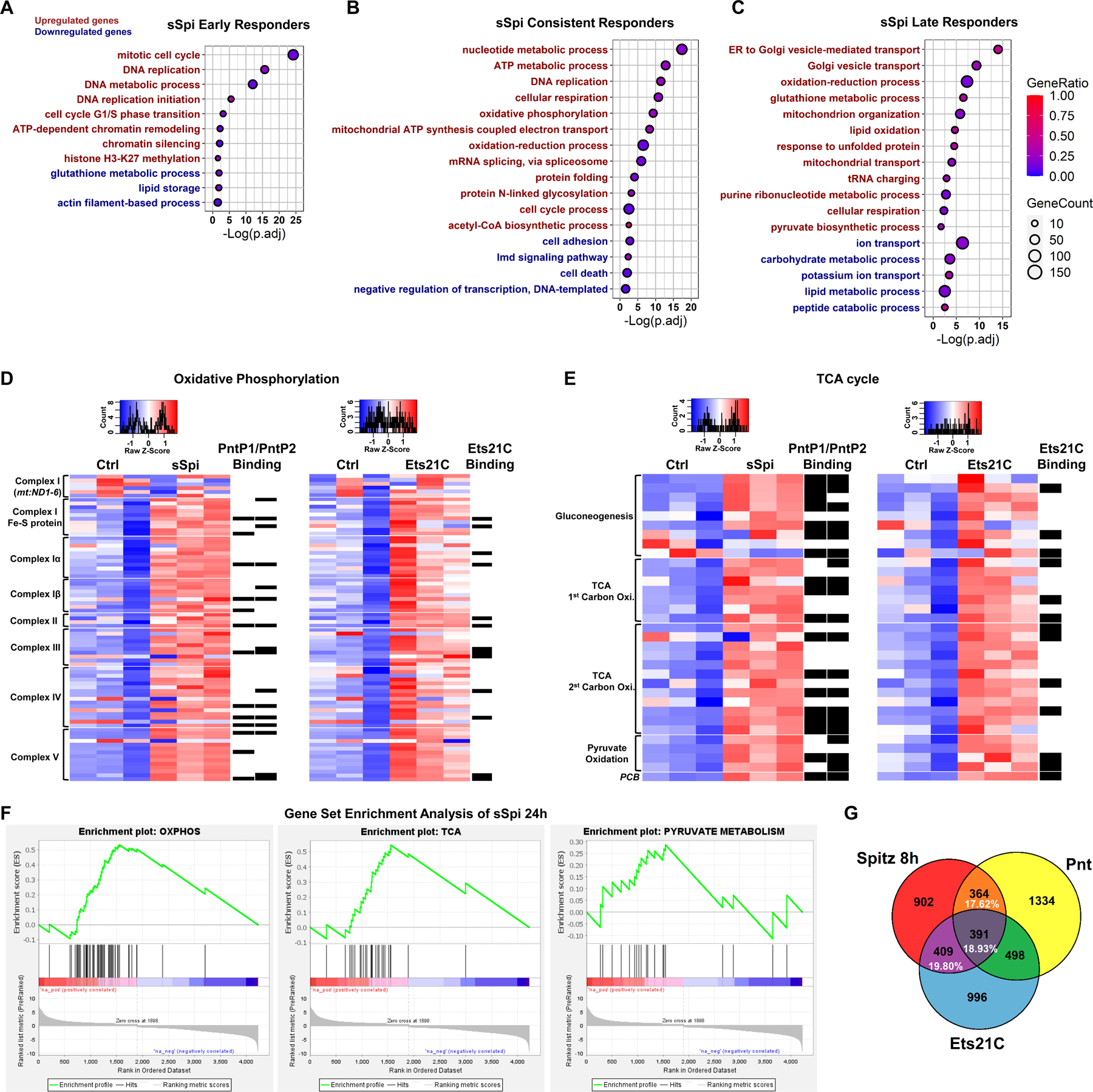
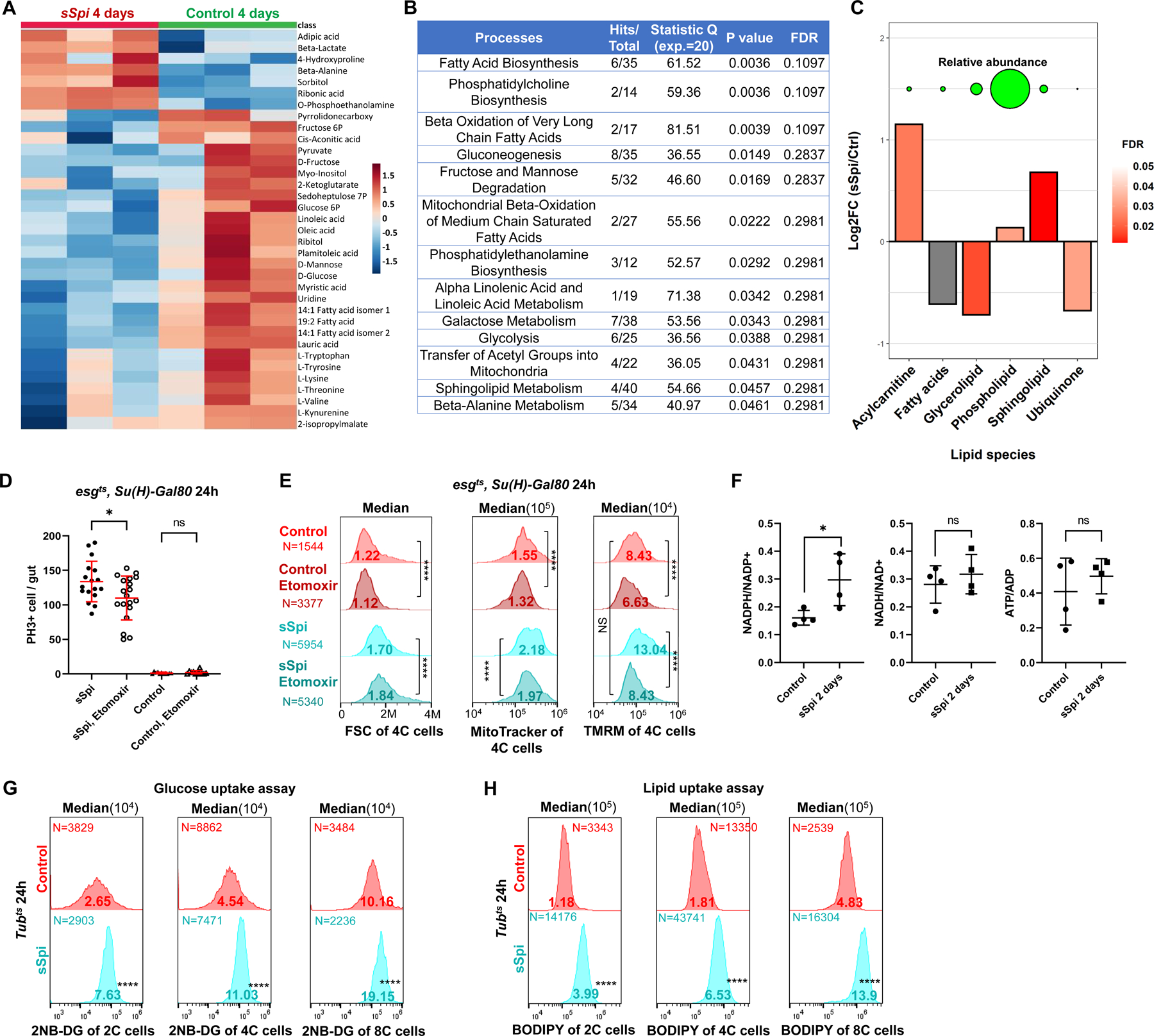
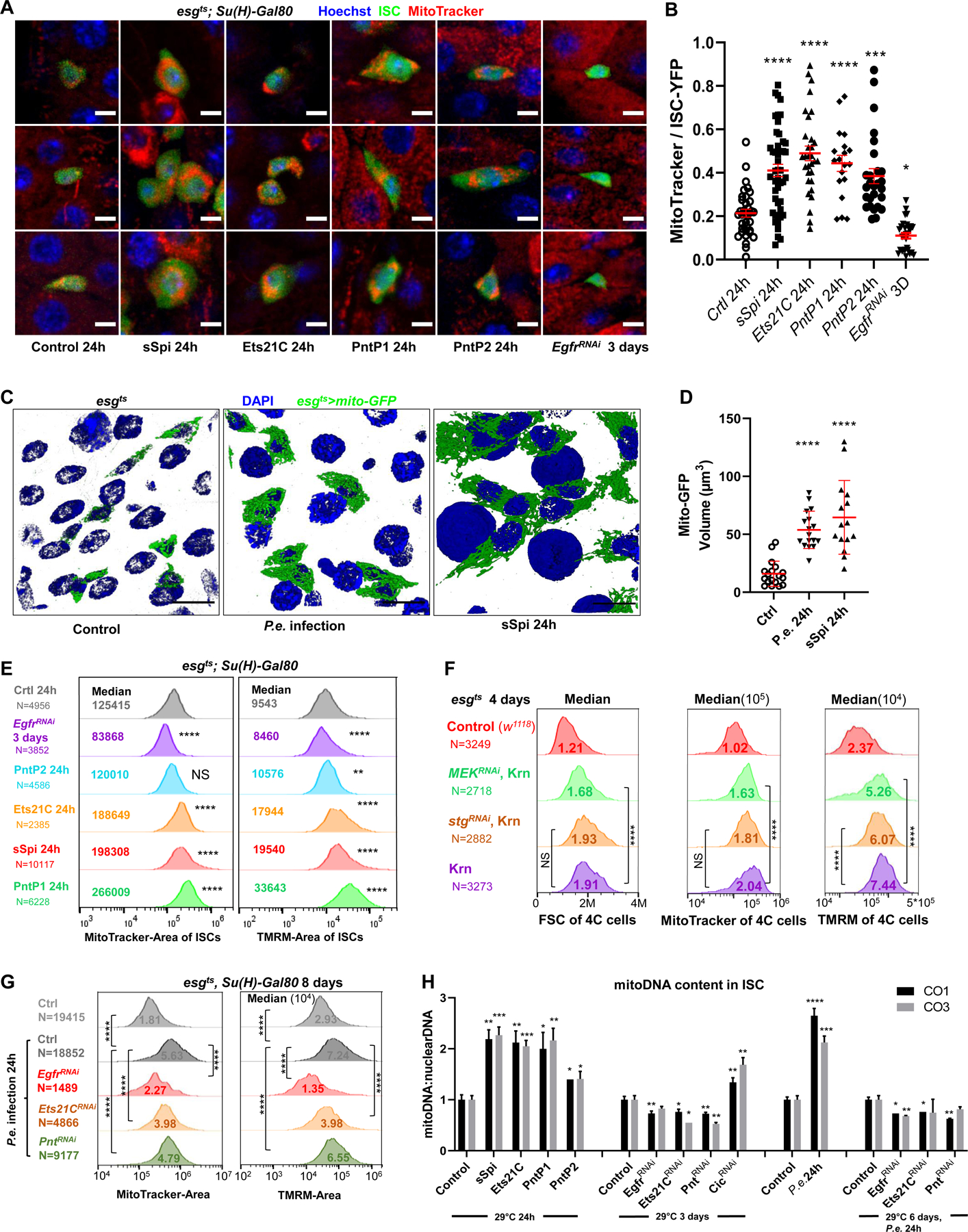
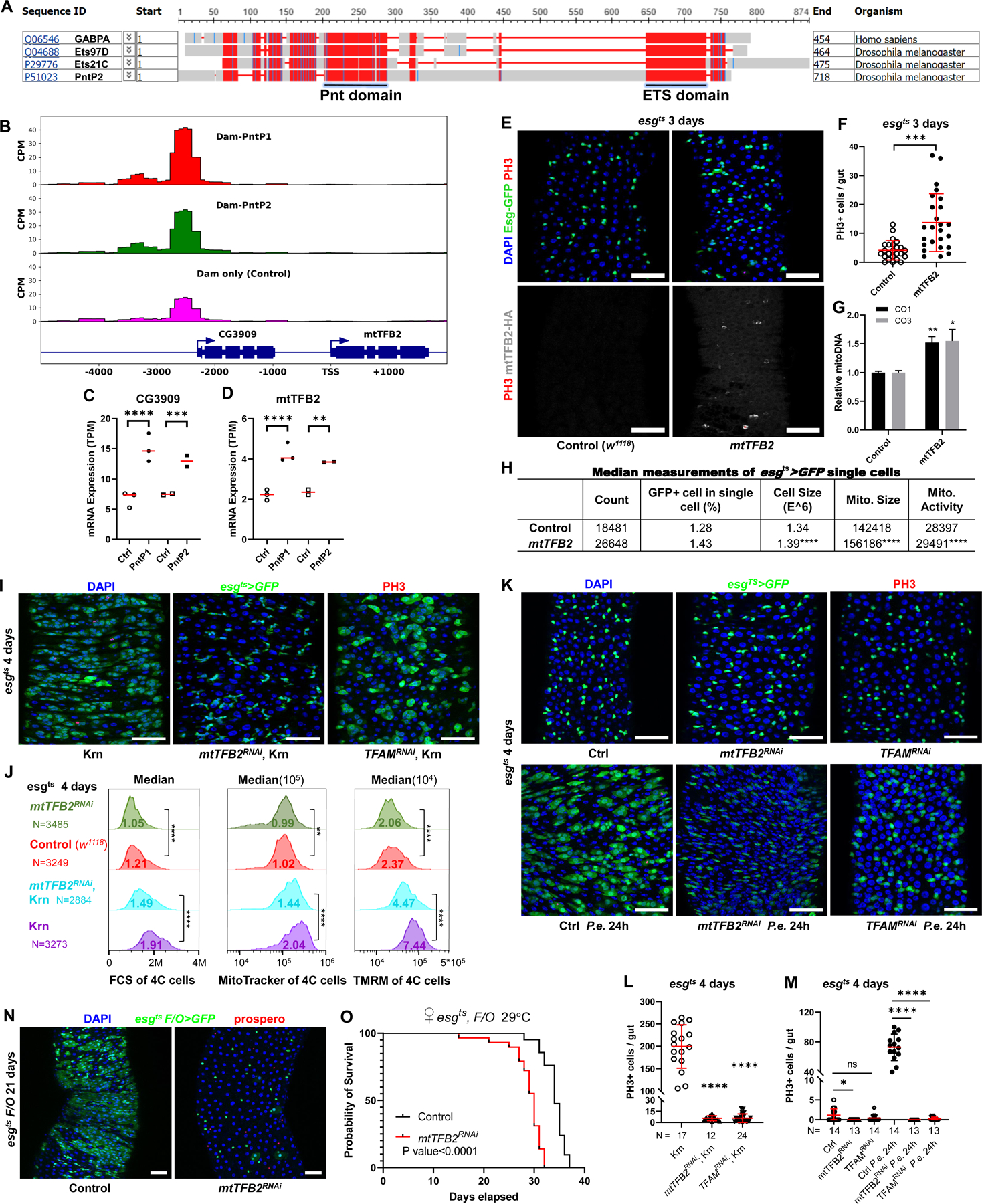
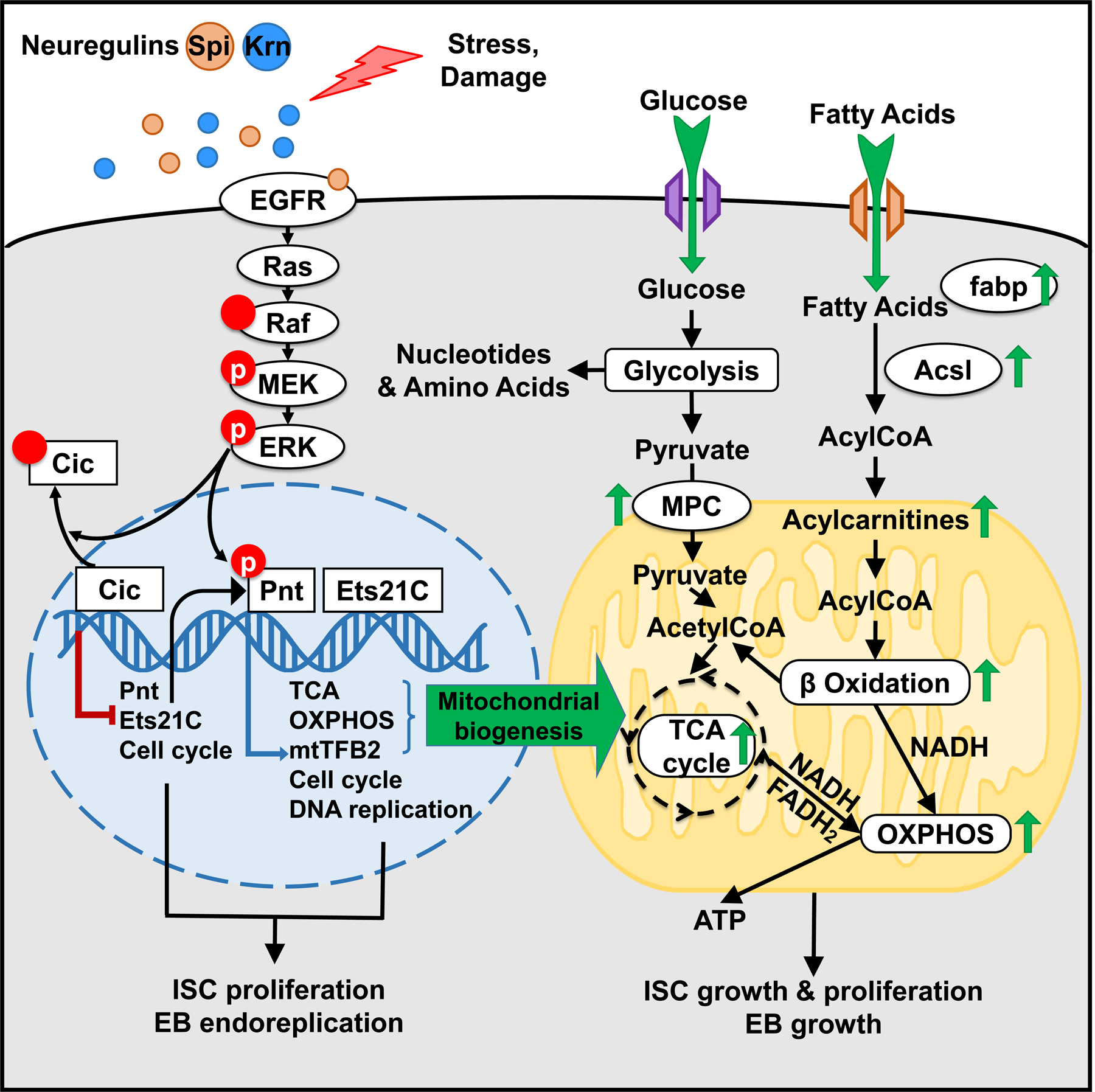
EGFR-RAS-ERK signaling promotes growth and proliferation in many cell types, and genetic hyperactivation of RAS-ERK signaling drives many cancers. Yet, despite intensive study of upstream components in EGFR signal transduction, the identities and functions of downstream effectors in the pathway are poorly understood. In Drosophila intestinal stem cells (ISCs), the transcriptional repressor Capicua (Cic) and its targets, the ETS-type transcriptional activators Pointed (pnt) and Ets21C, are essential downstream effectors of mitogenic EGFR signaling. Here, we show that these factors promote EGFR-dependent metabolic changes that increase ISC mass, mitochondrial growth, and mitochondrial activity. Gene target analysis using RNA and DamID sequencing revealed that Pnt and Ets21C directly upregulate not only DNA replication and cell cycle genes but also genes for oxidative phosphorylation, the TCA cycle, and fatty acid beta-oxidation. Metabolite analysis substantiated these metabolic functions. The mitochondrial transcription factor B2 (mtTFB2), a direct target of Pnt, was required and partially sufficient for EGFR-driven ISC growth, mitochondrial biogenesis, and proliferation. MEK-dependent EGF signaling stimulated mitochondrial biogenesis in human RPE-1 cells, indicating the conservation of these metabolic effects. This work illustrates how EGFR signaling alters metabolism to coordinately activate cell growth and cell division.
Keywords: Ets21C; ISC; Pointed; intestinal stem cell; mitochondrial biogenesis; mtTFB2; proliferation.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Stem cells: Mitochondrial biogenesis links growth and EGFR signaling.Curr Biol. 2022 Sep 12;32(17):R931-R934. doi: 10.1016/j.cub.2022.08.002. Curr Biol. 2022. PMID: 36099901
References
-
- Xu N, Wang SQ, Tan D, Gao Y, Lin G, and Xi R (2011). EGFR, Wingless and JAK/STAT signaling cooperatively maintain Drosophila intestinal stem cells. Developmental Biology 354, 31–43. - PubMed
-
- Basak O, Beumer J, Wiebrands K, Seno H, van Oudenaarden A, and Clevers H (2017). Induced Quiescence of Lgr5+ Stem Cells in Intestinal Organoids Enables Differentiation of Hormone-Producing Enteroendocrine Cells. Cell Stem Cell 20, 177–190.e4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
