

The LAMMER Kinase MoKns1 Regulates Growth, Conidiation and Pathogenicity in Magnaporthe oryzae
- PMID: 35897680
- PMCID: PMC9332457
- DOI: 10.3390/ijms23158104
The LAMMER Kinase MoKns1 Regulates Growth, Conidiation and Pathogenicity in Magnaporthe oryzae
Abstract
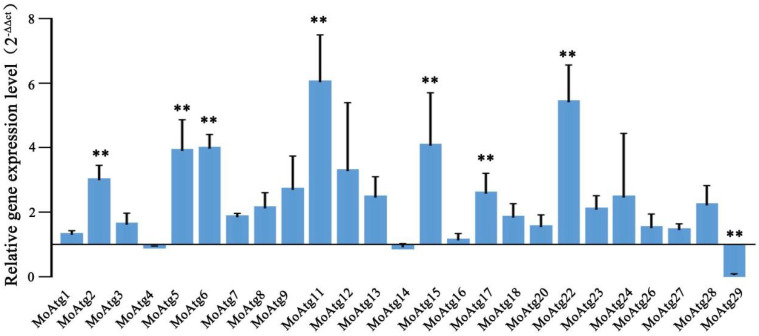
Magnaporthe oryzae is an important pathogen that causes a devastating disease in rice. It has been reported that the dual-specificity LAMMER kinase is conserved from yeast to animal species and has a variety of functions. However, the functions of the LAMMER kinase have not been reported in M. oryzae. In this study, we identified the unique LAMMER kinase MoKns1 and analyzed its function in M. oryzae. We found that in a MoKNS1 deletion mutant, growth and conidiation were primarily decreased, and pathogenicity was almost completely lost. Furthermore, our results found that MoKns1 is involved in autophagy. The ΔMokns1 mutant was sensitive to rapamycin, and MoKns1 interacted with the autophagy-related protein MoAtg18. Compared with the wild-type strain 70-15, autophagy was significantly enhanced in the ΔMokns1 mutant. In addition, we also found that MoKns1 regulated DNA damage stress pathways, and the ΔMokns1 mutant was more sensitive to hydroxyurea (HU) and methyl methanesulfonate (MMS) compared to the wild-type strain 70-15. The expression of genes related to DNA damage stress pathways in the ΔMokns1 mutant was significantly different from that in the wild-type strain. Our results demonstrate that MoKns1 is an important pathogenic factor in M. oryzae involved in regulating autophagy and DNA damage response pathways, thus affecting virulence. This research on M. oryzae pathogenesis lays a foundation for the prevention and control of M. oryzae.
Keywords: LAMMER kinase; Magnaporthe oryzae; conidiation; pathogenicity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
-
- Kong L.A., Li G.T., Liu Y., Liu M.G., Zhang S.J., Yang J., Zhou X.Y., Peng Y.L., Xu J.R. Differences between appressoria formed by germ tubes and appressorium-like structures developed by hyphal tips in Magnaporthe oryzae. Fungal Genet. Biol. 2013;56:33–41. doi: 10.1016/j.fgb.2013.03.006. - DOI - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
