

Palmitic acid control of ciliogenesis modulates insulin signaling in hypothalamic neurons through an autophagy-dependent mechanism
- PMID: 35902579
- PMCID: PMC9334645
- DOI: 10.1038/s41419-022-05109-9
Palmitic acid control of ciliogenesis modulates insulin signaling in hypothalamic neurons through an autophagy-dependent mechanism
Abstract
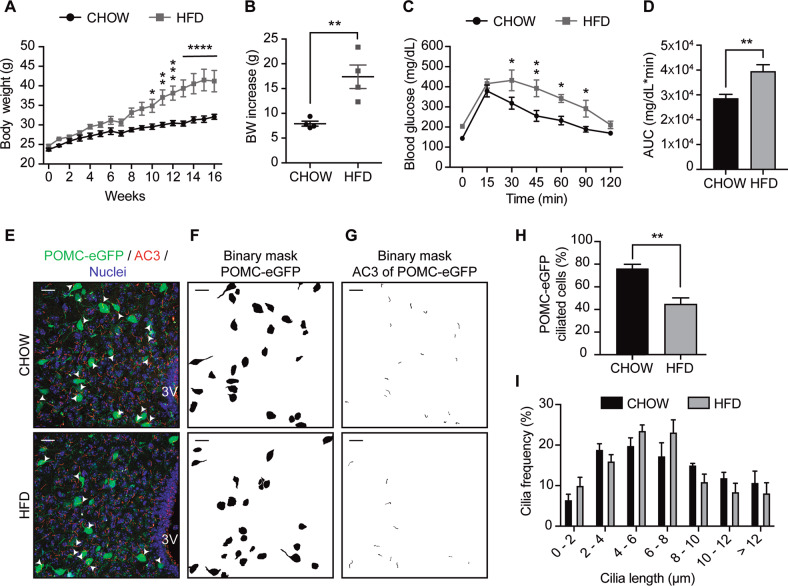
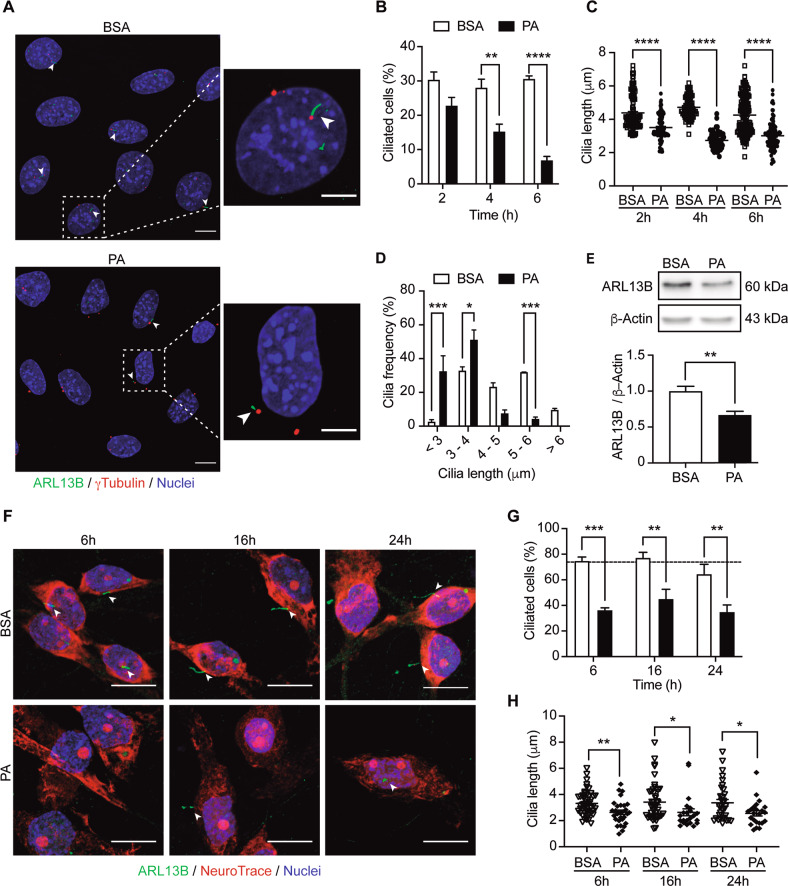
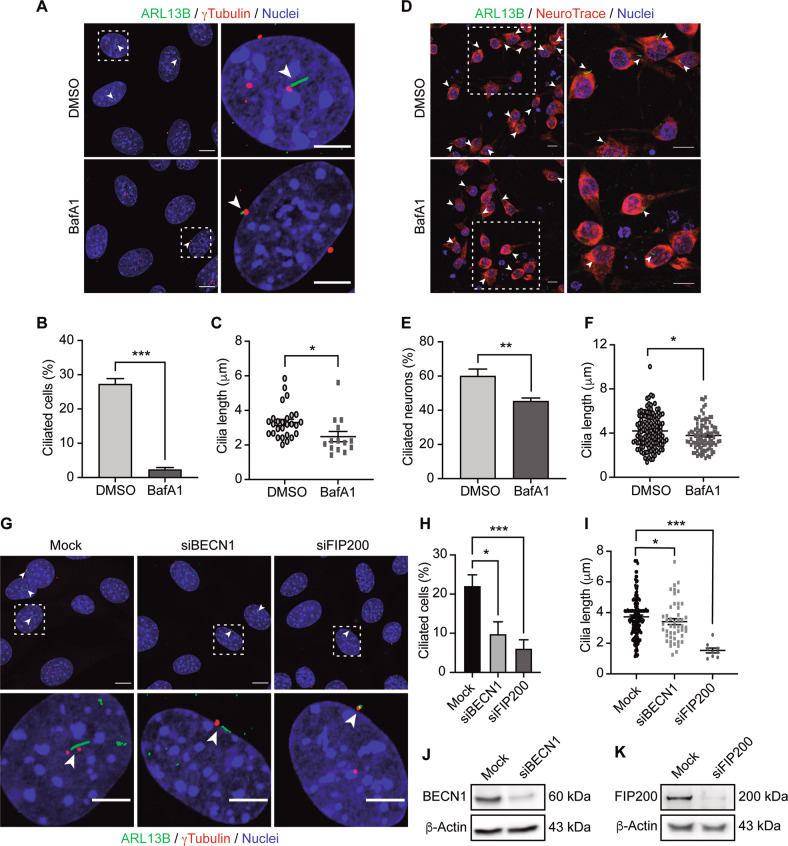
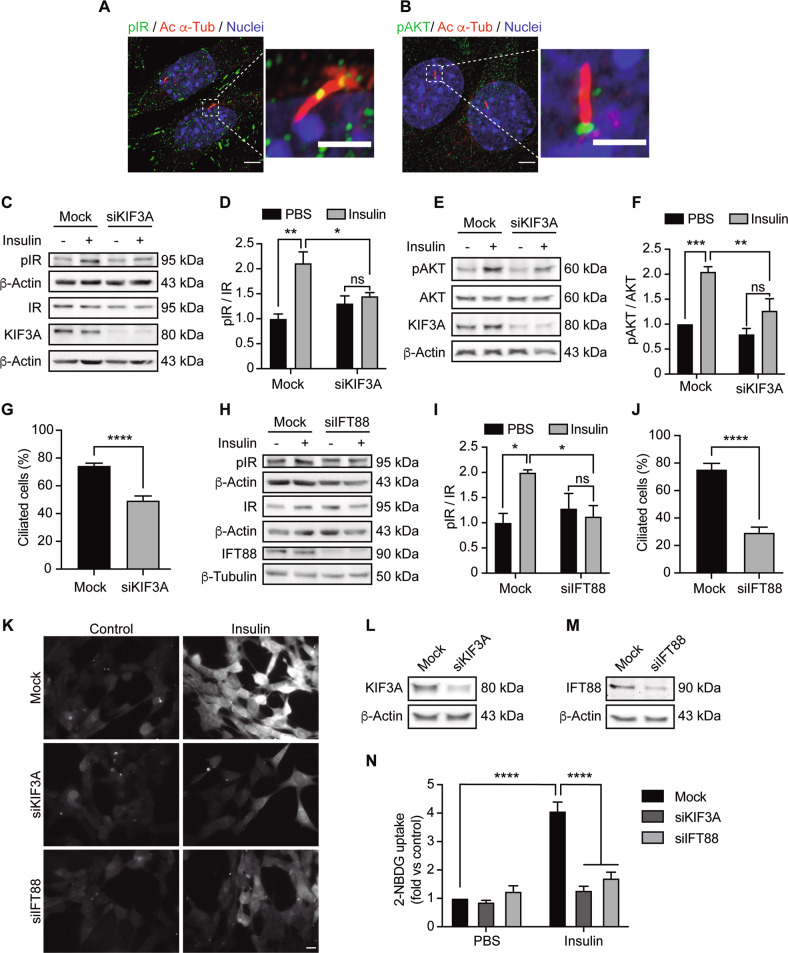
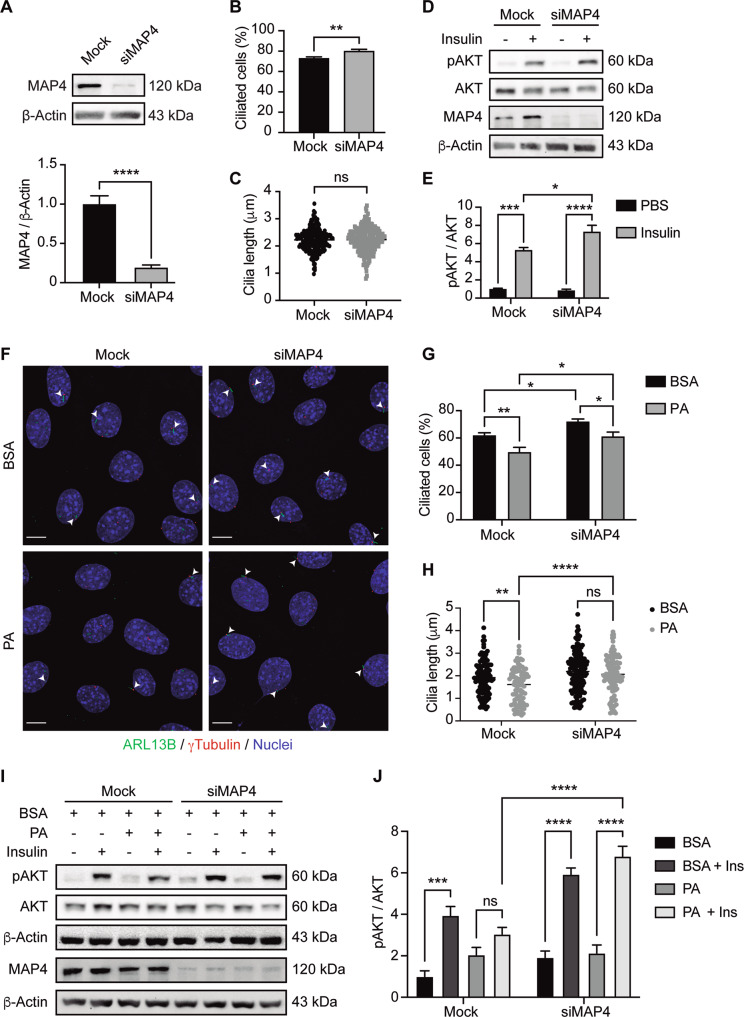
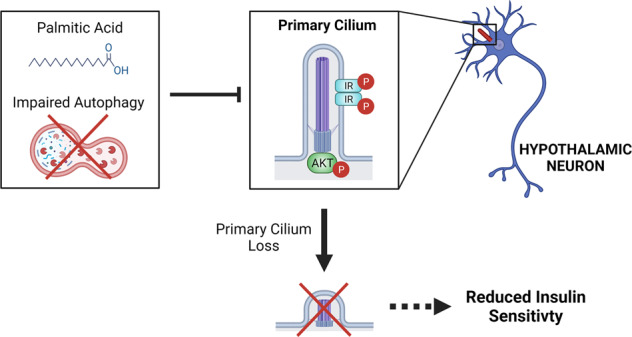
Palmitic acid (PA) is significantly increased in the hypothalamus of mice, when fed chronically with a high-fat diet (HFD). PA impairs insulin signaling in hypothalamic neurons, by a mechanism dependent on autophagy, a process of lysosomal-mediated degradation of cytoplasmic material. In addition, previous work shows a crosstalk between autophagy and the primary cilium (hereafter cilium), an antenna-like structure on the cell surface that acts as a signaling platform for the cell. Ciliopathies, human diseases characterized by cilia dysfunction, manifest, type 2 diabetes, among other features, suggesting a role of the cilium in insulin signaling. Cilium depletion in hypothalamic pro-opiomelanocortin (POMC) neurons triggers obesity and insulin resistance in mice, the same phenotype as mice deficient in autophagy in POMC neurons. Here we investigated the effect of chronic consumption of HFD on cilia; and our results indicate that chronic feeding with HFD reduces the percentage of cilia in hypothalamic POMC neurons. This effect may be due to an increased amount of PA, as treatment with this saturated fatty acid in vitro reduces the percentage of ciliated cells and cilia length in hypothalamic neurons. Importantly, the same effect of cilia depletion was obtained following chemical and genetic inhibition of autophagy, indicating autophagy is required for ciliogenesis. We further demonstrate a role for the cilium in insulin sensitivity, as cilium loss in hypothalamic neuronal cells disrupts insulin signaling and insulin-dependent glucose uptake, an effect that correlates with the ciliary localization of the insulin receptor (IR). Consistently, increased percentage of ciliated hypothalamic neuronal cells promotes insulin signaling, even when cells are exposed to PA. Altogether, our results indicate that, in hypothalamic neurons, impairment of autophagy, either by PA exposure, chemical or genetic manipulation, cause cilia loss that impairs insulin sensitivity.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Ng M, Fleming T, Robinson M, Thomson B, Graetz N, Margono C, et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: a systematic analysis for the Global Burden of Disease Study 2013. Lancet. 2014;384:766–81. doi: 10.1016/S0140-6736(14)60460-8. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
