

Phosphatidylinositol-4-phosphate controls autophagosome formation in Arabidopsis thaliana
- PMID: 35902598
- PMCID: PMC9334301
- DOI: 10.1038/s41467-022-32109-2
Phosphatidylinositol-4-phosphate controls autophagosome formation in Arabidopsis thaliana
Abstract
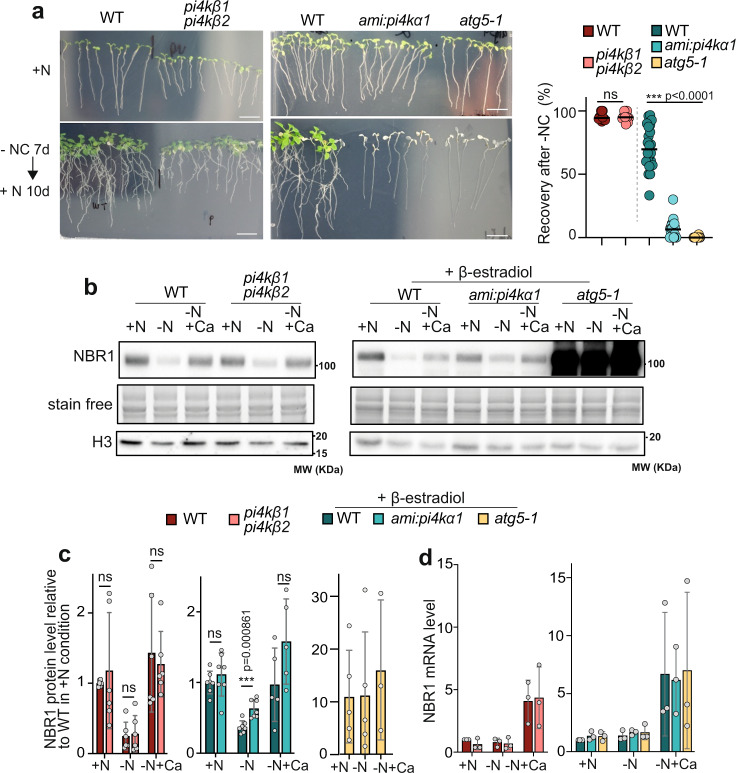
Autophagy is an intracellular degradation mechanism critical for plant acclimation to environmental stresses. Central to autophagy is the formation of specialized vesicles, the autophagosomes, which target and deliver cargo to the lytic vacuole. How autophagosomes form in plant cells remains poorly understood. Here, we uncover the importance of the lipid phosphatidylinositol-4-phosphate in autophagy using pharmacological and genetical approaches. Combining biochemical and live-microscopy analyses, we show that PI4K activity is required for early stages of autophagosome formation. Further, our results show that the plasma membrane-localized PI4Kα1 is involved in autophagy and that a substantial portion of autophagy structures are found in proximity to the PI4P-enriched plasma membrane. Together, our study unravels critical insights into the molecular determinants of autophagy, proposing a model whereby the plasma membrane provides PI4P to support the proper assembly and expansion of the phagophore thus governing autophagosome formation in Arabidopsis.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
