

UHRF1 interacts with snRNAs and regulates alternative splicing in mouse spermatogonial stem cells
- PMID: 35905740
- PMCID: PMC9391524
- DOI: 10.1016/j.stemcr.2022.06.010
UHRF1 interacts with snRNAs and regulates alternative splicing in mouse spermatogonial stem cells
Abstract
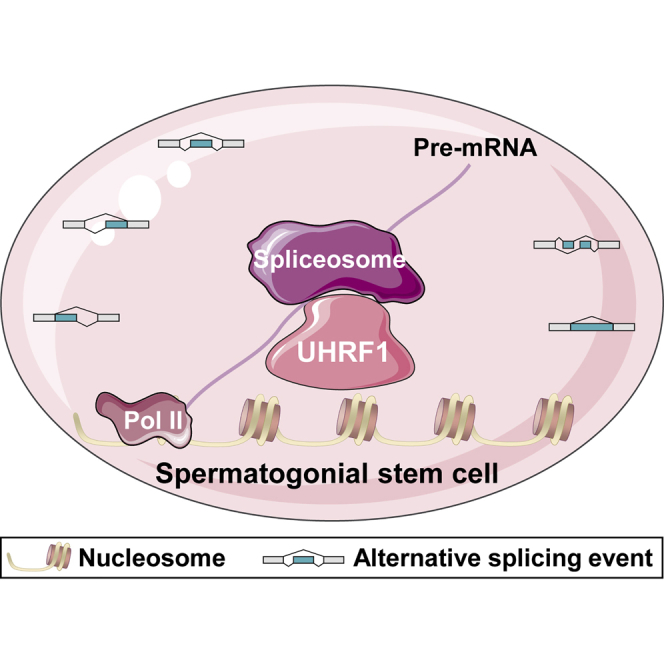
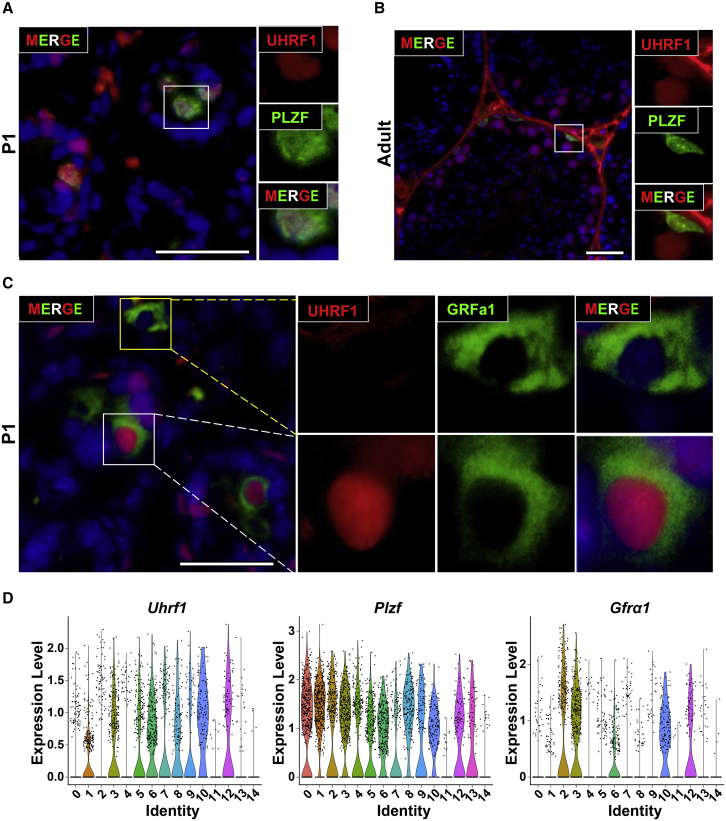
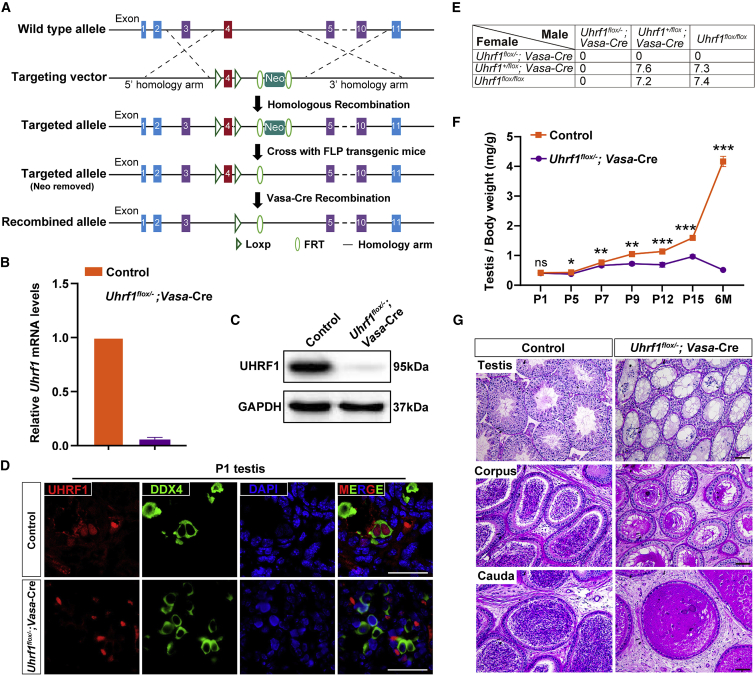
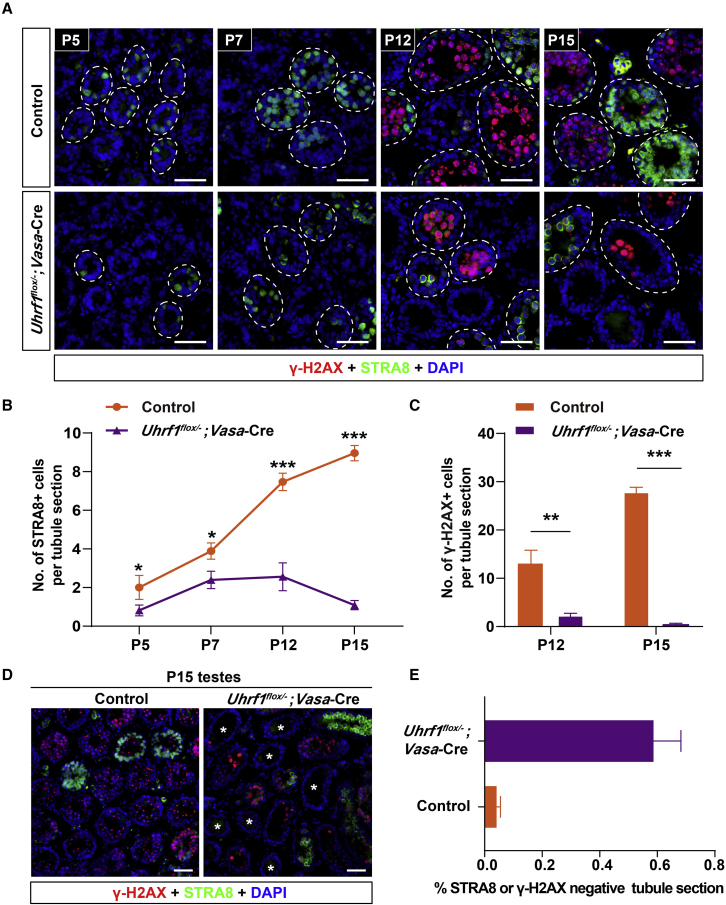
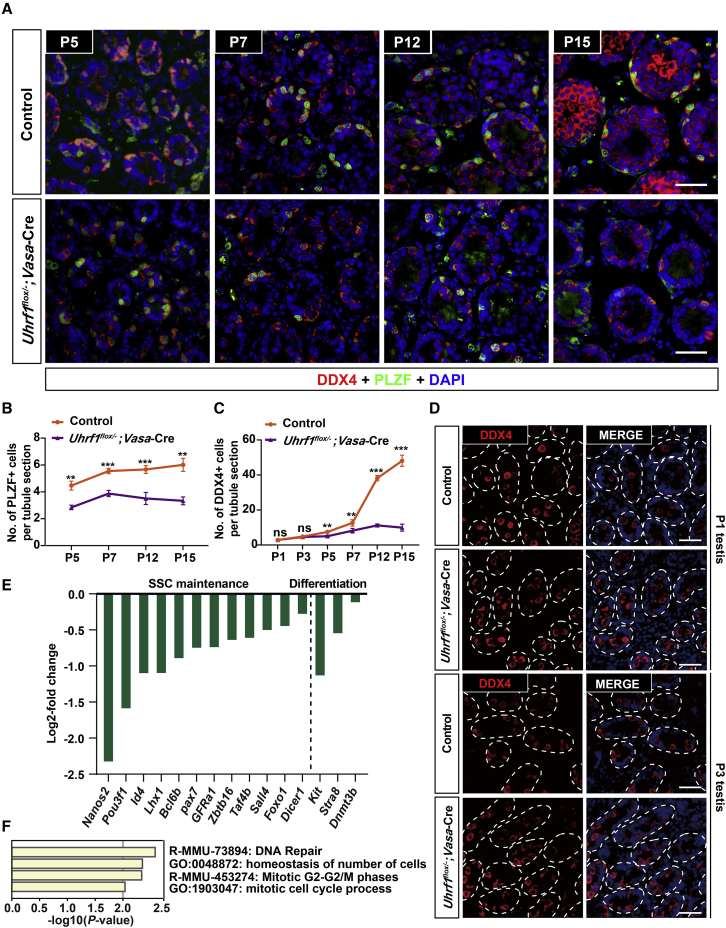
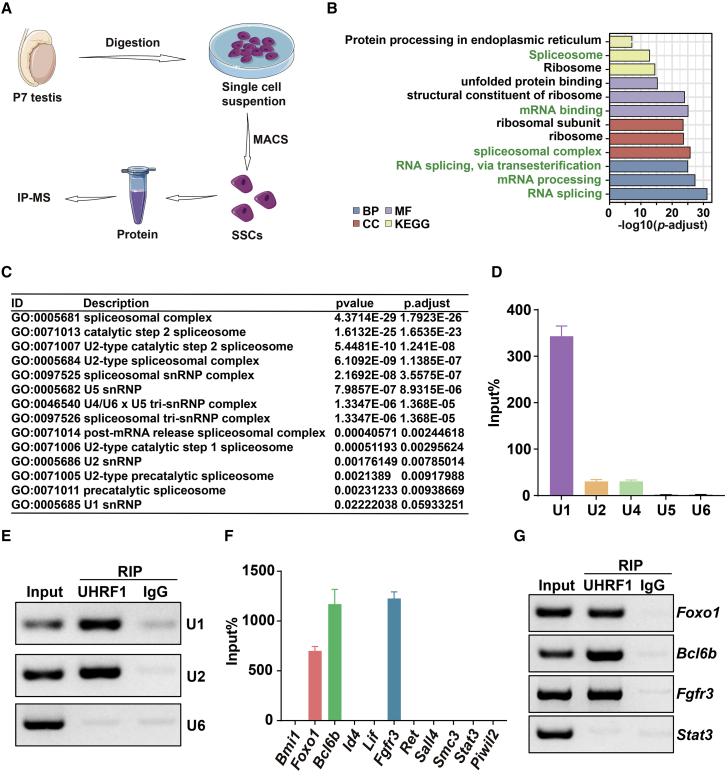
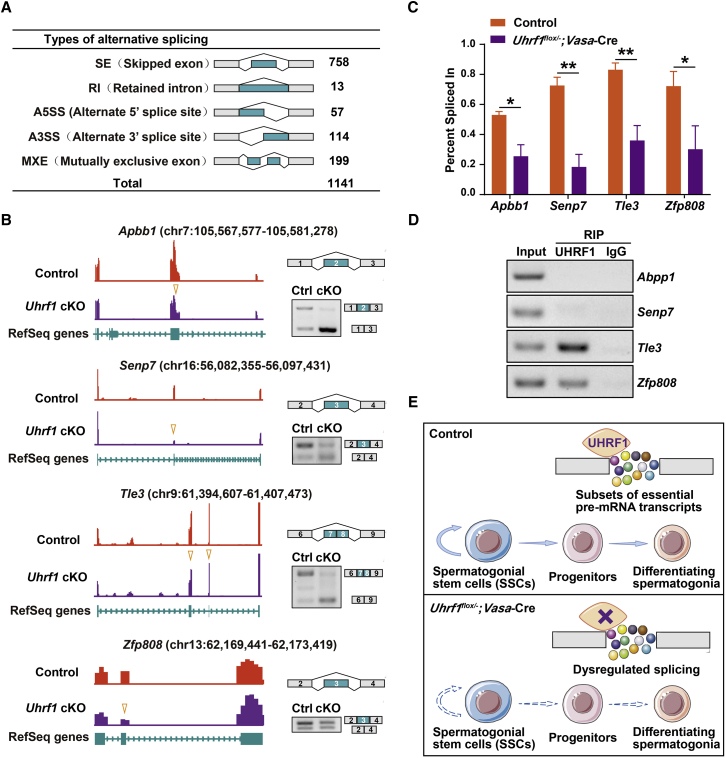
Life-long male fertility relies on exquisite homeostasis and the development of spermatogonial stem cells (SSCs); however, the underlying molecular genetic and epigenetic regulation in this equilibrium process remains unclear. Here, we document that UHRF1 interacts with snRNAs to regulate pre-mRNA alternative splicing in SSCs and is required for the homeostasis of SSCs in mice. Genetic deficiency of UHRF1 in mouse prospermatogonia results in gradual loss of spermatogonial stem cells, eventually leading to Sertoli-cell-only syndrome (SCOS) and male infertility. Comparative RNA-seq data provide evidence that Uhrf1 ablation dysregulates previously reported SSC maintenance- and differentiation-related genes. We further found that UHRF1 could act as an alternative RNA splicing regulator and interact with Tle3 transcripts to regulate its splicing event in spermatogonia. Collectively, our data reveal a multifunctional role for UHRF1 in regulating gene expression programs and alternative splicing during SSC homeostasis, which may provide clues for treating human male infertility.
Keywords: Sertoli-cell-only syndrome; UHRF1; alternative splicing; snRNA; spermatogenesis; spermatogonial stem cell.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interests The authors declare no competing interests.
Figures
References
-
- Achour M., Jacq X., Rondé P., Alhosin M., Charlot C., Chataigneau T., Jeanblanc M., Macaluso M., Giordano A., Hughes A.D., et al. The interaction of the SRA domain of ICBP90 with a novel domain of DNMT1 is involved in the regulation of VEGF gene expression. Oncogene. 2008;27:2187–2197. doi: 10.1038/sj.onc.1210855. - DOI - PubMed
-
- Arita K., Isogai S., Oda T., Unoki M., Sugita K., Sekiyama N., Kuwata K., Hamamoto R., Tochio H., Sato M., et al. Recognition of modification status on a histone H3 tail by linked histone reader modules of the epigenetic regulator UHRF1. Proc. Natl. Acad. Sci. USA. 2012;109:12950–12955. doi: 10.1073/pnas.1203701109. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
