

A novel mouse model of CMT1B identifies hyperglycosylation as a new pathogenetic mechanism
- PMID: 35908287
- PMCID: PMC9759335
- DOI: 10.1093/hmg/ddac170
A novel mouse model of CMT1B identifies hyperglycosylation as a new pathogenetic mechanism
Abstract
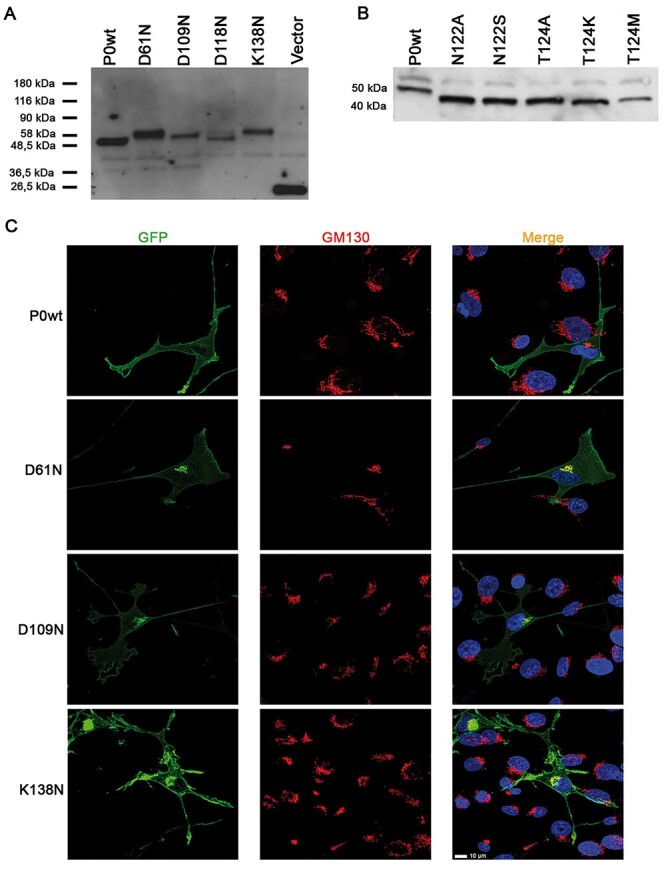
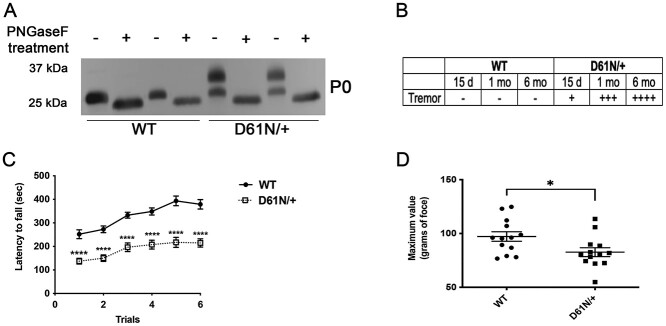
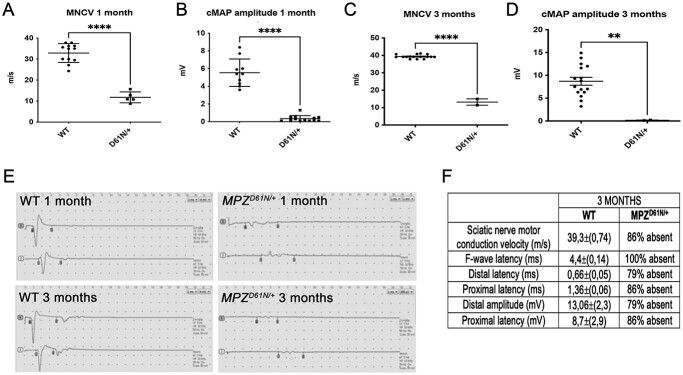
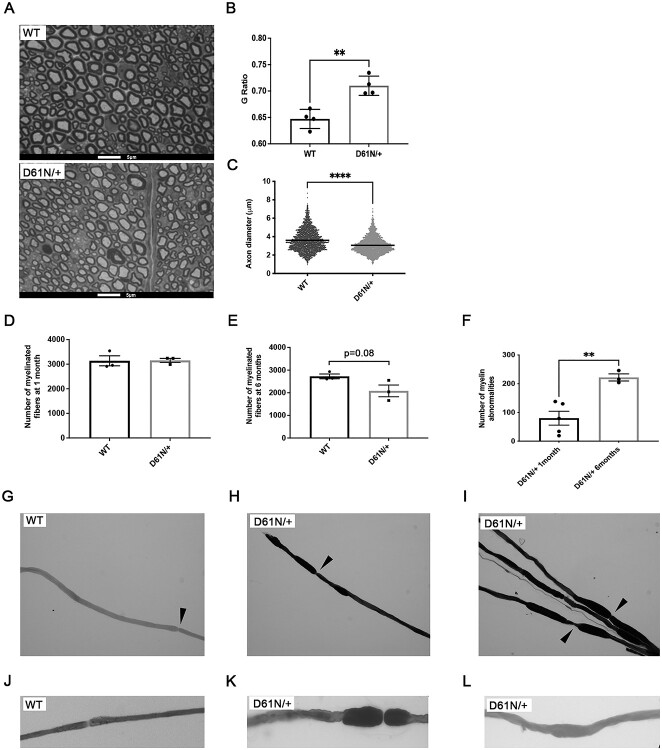
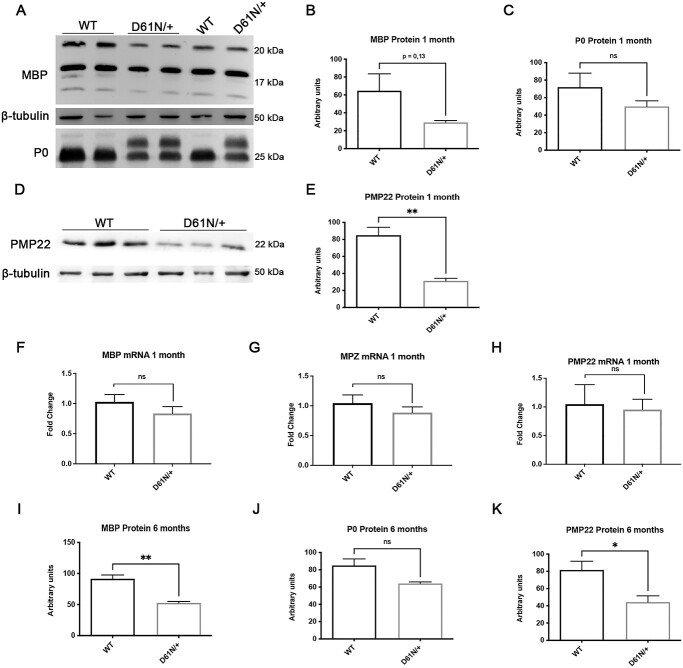
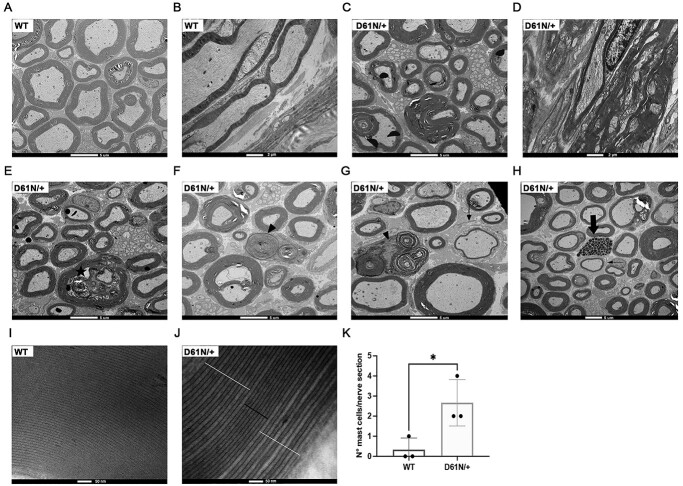
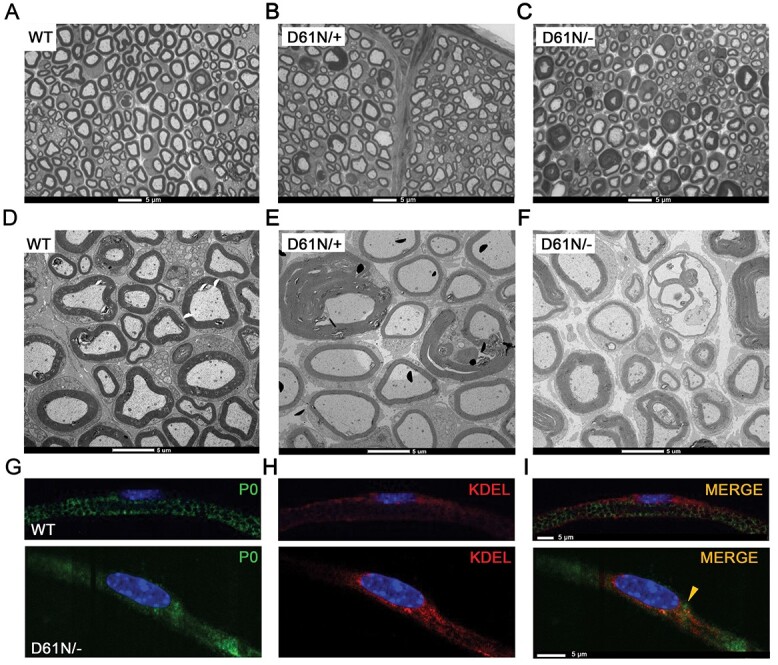
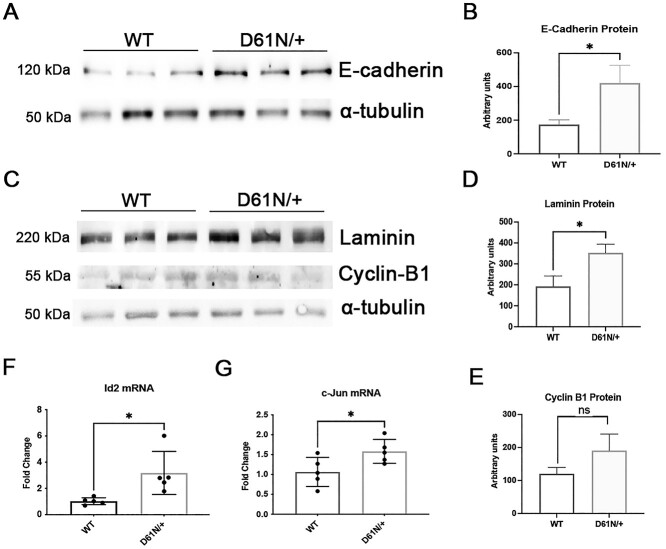
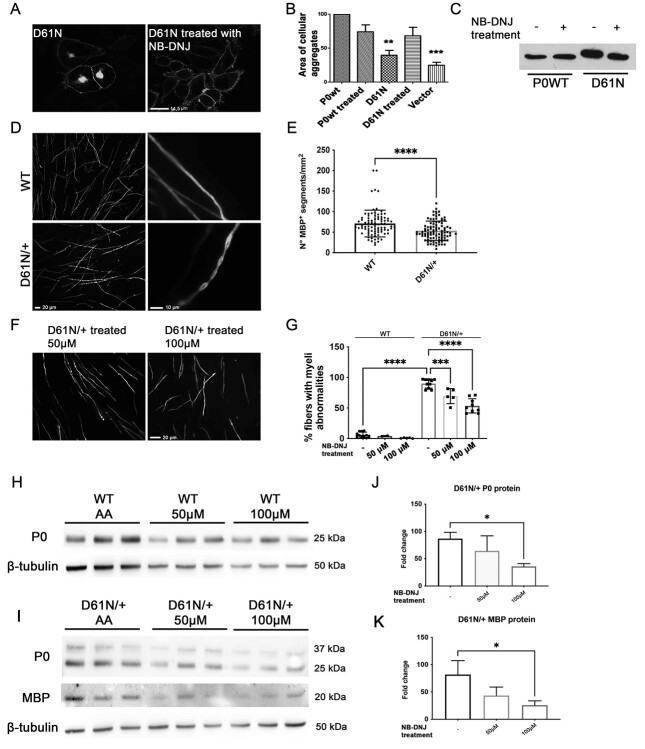
Mutations in the Myelin Protein Zero gene (MPZ), encoding P0, the major structural glycoprotein of peripheral nerve myelin, are the cause of Charcot-Marie-Tooth (CMT) type 1B neuropathy, and most P0 mutations appear to act through gain-of-function mechanisms. Here, we investigated how misglycosylation, a pathomechanism encompassing several genetic disorders, may affect P0 function. Using in vitro assays, we showed that gain of glycosylation is more damaging for P0 trafficking and functionality as compared with a loss of glycosylation. Hence, we generated, via CRISPR/Cas9, a mouse model carrying the MPZD61N mutation, predicted to generate a new N-glycosylation site in P0. In humans, MPZD61N causes a severe early-onset form of CMT1B, suggesting that hyperglycosylation may interfere with myelin formation, leading to pathology. We show here that MPZD61N/+ mice develop a tremor as early as P15 which worsens with age and correlates with a significant motor impairment, reduced muscular strength and substantial alterations in neurophysiology. The pathological analysis confirmed a dysmyelinating phenotype characterized by diffuse hypomyelination and focal hypermyelination. We find that the mutant P0D61N does not cause significant endoplasmic reticulum stress, a common pathomechanism in CMT1B, but is properly trafficked to myelin where it causes myelin uncompaction. Finally, we show that myelinating dorsal root ganglia cultures from MPZD61N mice replicate some of the abnormalities seen in vivo, suggesting that they may represent a valuable tool to investigate therapeutic approaches. Collectively, our data indicate that the MPZD61N/+ mouse represents an authentic model of severe CMT1B affirming gain-of-glycosylation in P0 as a novel pathomechanism of disease.
© The Author(s) 2022. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Sherman, D.L. and Brophy, P.J. (2005) Mechanisms of axon ensheathment and myelin growth. Nat. Rev. Neurosci., 6, 683–690. - PubMed
-
- Jessen, K.R. and Mirsky, R. (2005) The origin and development of glial cells in peripheral nerves. Nat. Rev. Neurosci., 6, 671–682. - PubMed
-
- Patzig, J., Jahn, O., Tenzer, S., Wichert, S.P., de Monasterio-Schrader, P., Rosfa, S., Kuharev, J., Yan, K., Bormuth, I., Bremer, J. et al. (2011) Quantitative and integrative proteome analysis of peripheral nerve myelin identifies novel myelin proteins and candidate neuropathy loci. J. Neurosci., 31, 16369–16386. - PMC - PubMed
-
- Filbin, M.T., Walsh, F.S., Trapp, B.D., Pizzey, J.A. and Tennekoon, G.I. (1990) Role of myelin P0 protein as a homophilic adhesion molecule. Nature, 344, 871–872. - PubMed
-
- Shapiro, L., Doyle, J.P., Hensley, P., Colman, D.R. and Hendrickson, W.A. (1996) Crystal structure of the extracellular domain from P0, the major structural protein of peripheral nerve myelin. Neuron, 17, 435–449. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
