

Chloroplastic photoprotective strategies differ between bundle sheath and mesophyll cells in maize (Zea mays L.) Under drought
- PMID: 35909748
- PMCID: PMC9330506
- DOI: 10.3389/fpls.2022.885781
Chloroplastic photoprotective strategies differ between bundle sheath and mesophyll cells in maize (Zea mays L.) Under drought
Abstract
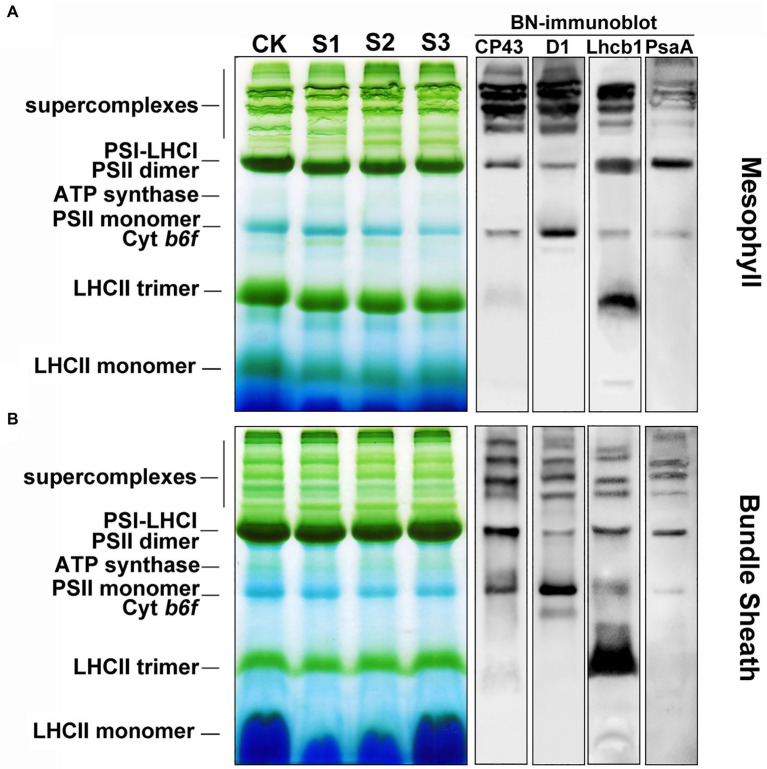
Bundle sheath cells play a crucial role in photosynthesis in C4 plants, but the structure and function of photosystem II (PSII) in these cells is still controversial. Photoprotective roles of bundle sheath chloroplasts at the occurrence of environmental stresses have not been investigated so far. Non-photochemical quenching (NPQ) of chlorophyll a fluorescence is the photoprotective mechanism that responds to a changing energy balance in chloroplasts. In the present study, we found a much higher NPQ in bundle sheath chloroplasts than in mesophyll chloroplasts under a drought stress. This change was accompanied by a more rapid dephosphorylation of light-harvesting complex II (LHCII) subunits and a greater increase in PSII subunit S (PsbS) protein abundance than in mesophyll cell chloroplasts. Histochemical staining of reactive oxygen species (ROS) suggested that the high NPQ may be one of the main reasons for the lower accumulation of ROS in bundle sheath chloroplasts. This may maintain the stable functioning of bundle sheath cells under drought condition. These results indicate that the superior capacity for dissipation of excitation energy in bundle sheath chloroplasts may be an environmental adaptation unique to C4 plants.
Keywords: bundle sheath chloroplast; drought stress; maize (Zea mays L.); non-photochemical quenching; photoprotection; reactive oxygen species.
Copyright © 2022 Liu, Liu, Chen, Yin, Zhang, Song, Chang, Zhang, Wang, Dai, Wei, Xiong, Yuan and Zhao.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures







Similar articles
-
Acclimation of mesophyll and bundle sheath chloroplasts of maize to different irradiances during growth.Biochim Biophys Acta. 2006 Nov;1757(11):1539-46. doi: 10.1016/j.bbabio.2006.09.001. Epub 2006 Sep 14. Biochim Biophys Acta. 2006. PMID: 17034754
-
Biochemical properties and ultrastructure of mesophyll and bundle sheath thylakoids from maize (Zea mays) chloroplasts.Funct Plant Biol. 2020 Oct;47(11):970-976. doi: 10.1071/FP20004. Funct Plant Biol. 2020. PMID: 32574552
-
Photochemical properties of mesophyll and bundle sheath chloroplasts of maize.Plant Physiol. 1973 Sep;52(3):257-62. doi: 10.1104/pp.52.3.257. Plant Physiol. 1973. PMID: 16658543 Free PMC article.
-
Differential positioning of chloroplasts in C4 mesophyll and bundle sheath cells.Plant Signal Behav. 2011 Aug;6(8):1111-3. doi: 10.4161/psb.6.8.15809. Epub 2011 Aug 1. Plant Signal Behav. 2011. PMID: 21757999 Free PMC article. Review.
-
Reactive oxygen species and redox regulation in mesophyll and bundle sheath cells of C4 plants.J Exp Bot. 2018 Jun 19;69(14):3321-3331. doi: 10.1093/jxb/ery064. J Exp Bot. 2018. PMID: 29529246 Review.
Cited by
-
Nanobiotechnology-mediated regulation of reactive oxygen species homeostasis under heat and drought stress in plants.Front Plant Sci. 2024 Aug 27;15:1418515. doi: 10.3389/fpls.2024.1418515. eCollection 2024. Front Plant Sci. 2024. PMID: 39258292 Free PMC article. Review.
-
Drought responsiveness in six wheat genotypes: identification of stress resistance indicators.Front Plant Sci. 2023 Sep 13;14:1232583. doi: 10.3389/fpls.2023.1232583. eCollection 2023. Front Plant Sci. 2023. PMID: 37780517 Free PMC article.
-
Differential regulation of reactive oxygen species in dimorphic chloroplasts of single cell C4 plant Bienertia sinuspersici during drought and salt stress.Front Plant Sci. 2023 Apr 20;14:1030413. doi: 10.3389/fpls.2023.1030413. eCollection 2023. Front Plant Sci. 2023. PMID: 37152138 Free PMC article.
-
Developing Functional Relationships between Soil Moisture Content and Corn Early-Season Physiology, Growth, and Development.Plants (Basel). 2023 Jun 28;12(13):2471. doi: 10.3390/plants12132471. Plants (Basel). 2023. PMID: 37447032 Free PMC article.
-
Tomato Biodiversity and Drought Tolerance: A Multilevel Review.Int J Mol Sci. 2023 Jun 12;24(12):10044. doi: 10.3390/ijms241210044. Int J Mol Sci. 2023. PMID: 37373193 Free PMC article. Review.
References
-
- Campos H., Trejob C., Peña-Valdivia C. B., García-Nava R., Conde-Martínez F. V., Cruz-Ortega M. R. (2014). Stomatal and non-stomatal limitations of bell pepper (Capsicum annuum L.) plants under water stress and re-watering: delayed restoration of photosynthesis during recovery. Environ. Exp. Bot. 98, 56–64. doi: 10.1016/j.envexpbot.2013.10.015 - DOI
LinkOut - more resources
Full Text Sources
Miscellaneous