

SlCESTA Is a Brassinosteroid-Regulated bHLH Transcription Factor of Tomato That Promotes Chilling Tolerance and Fruit Growth When Over-Expressed
- PMID: 35909777
- PMCID: PMC9337221
- DOI: 10.3389/fpls.2022.930805
SlCESTA Is a Brassinosteroid-Regulated bHLH Transcription Factor of Tomato That Promotes Chilling Tolerance and Fruit Growth When Over-Expressed
Abstract
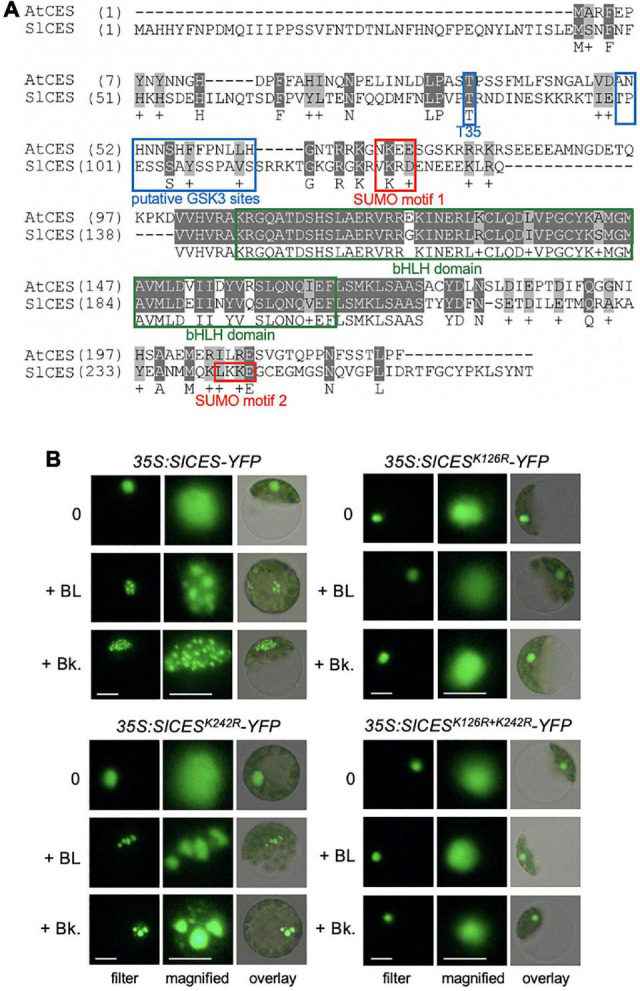
Brassinosteroids (BRs) are required for various aspects of plant growth and development, but also participate in stress responses. The hormones convey their activity through transcriptional regulation and posttranslational modification of transcription factors and one class are basic helix-loop-helix (bHLH) proteins of the BR Enhanced Expression (BEE) subfamily, which in Arabidopsis thaliana include BEE1-3 and CESTA (CES). CES and the BEEs promote the expression of different BR-responsive genes, including genes encoding gibberellin (GA) biosynthetic and catabolizing enzymes, as well as cold-responsive genes. Interestingly, in terms of an application, CES could promote both fruit growth and cold stress tolerance when over-expressed in A. thaliana and here it was investigated, if this function is conserved in the fruit crop Solanum lycopersicum (cultivated tomato). Based on amino acid sequence similarity and the presence of regulatory motifs, a CES orthologue of S. lycopersicum, SlCES, was identified and the effects of its over-expression were analysed in tomato. This showed that SlCES, like AtCES, was re-localized to nuclear bodies in response to BR signaling activation and that it effected GA homeostasis, with related phenotypes, when over-expressed. In addition, over-expression lines showed an increased chilling tolerance and had altered fruit characteristics. The possibilities and potential limitations of a gain of SlCES function as a breeding strategy for tomato are discussed.
Keywords: BEEs; SUMOylation; Solanum; brassinosteroids; gibberellins; hormones.
Copyright © 2022 Shuai, Chen, Wlk, Rozhon, Pimenta Lange, Sieberer, Lange and Poppenberger.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



Similar articles
-
Brassinosteroid-regulated bHLH transcription factor CESTA induces the gibberellin 2-oxidase GA2ox7.Plant Physiol. 2022 Mar 28;188(4):2012-2025. doi: 10.1093/plphys/kiac008. Plant Physiol. 2022. PMID: 35148416 Free PMC article.
-
Brassinosteroids participate in the control of basal and acquired freezing tolerance of plants.Proc Natl Acad Sci U S A. 2016 Oct 4;113(40):E5982-E5991. doi: 10.1073/pnas.1611477113. Epub 2016 Sep 21. Proc Natl Acad Sci U S A. 2016. PMID: 27655893 Free PMC article.
-
Enhanced brassinosteroid signaling via the overexpression of SlBRI1 positively regulates the chilling stress tolerance of tomato.Plant Sci. 2022 Jul;320:111281. doi: 10.1016/j.plantsci.2022.111281. Epub 2022 Apr 4. Plant Sci. 2022. PMID: 35643607
-
Interplay between phosphorylation and SUMOylation events determines CESTA protein fate in brassinosteroid signalling.Nat Commun. 2014 Aug 19;5:4687. doi: 10.1038/ncomms5687. Nat Commun. 2014. PMID: 25134617 Free PMC article.
-
Three redundant brassinosteroid early response genes encode putative bHLH transcription factors required for normal growth.Genetics. 2002 Nov;162(3):1445-56. doi: 10.1093/genetics/162.3.1445. Genetics. 2002. PMID: 12454087 Free PMC article.
Cited by
-
SlUPA-like, a bHLH Transcription Factor in Tomato (Solanum lycopersicum), Serves as the Crosstalk of GA, JA and BR.Int J Mol Sci. 2024 Dec 14;25(24):13419. doi: 10.3390/ijms252413419. Int J Mol Sci. 2024. PMID: 39769191 Free PMC article.
-
Genome-Wide Identification and Characterization of the bHLH Gene Family and Its Response to Abiotic Stresses in Carthamus tinctorius.Plants (Basel). 2023 Nov 3;12(21):3764. doi: 10.3390/plants12213764. Plants (Basel). 2023. PMID: 37960120 Free PMC article.
-
Genome- and Transcriptome-Wide Characterization and Expression Analyses of bHLH Transcription Factor Family Reveal Their Relevance to Salt Stress Response in Tomato.Plants (Basel). 2025 Jan 12;14(2):200. doi: 10.3390/plants14020200. Plants (Basel). 2025. PMID: 39861553 Free PMC article.
-
Manipulating brassinosteroid signaling pathway to genetically improve horticultural plants.aBIOTECH. 2025 Feb 22;6(2):328-345. doi: 10.1007/s42994-025-00201-y. eCollection 2025 Jun. aBIOTECH. 2025. PMID: 40641636 Free PMC article. Review.
References
-
- Achard P., Gong F., Cheminant S., Alioua M., Hedden P., Genschik P. (2008). The cold-inducible CBF1 factor-dependent signaling pathway modulates the accumulation of the growth-repressing DELLA proteins via its effect on gibberellin metabolism. Plant Cell 20 2117–2129. 10.1105/tpc.108.058941 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources