

Exogenous proline enhances susceptibility of NSCLC to cisplatin via metabolic reprogramming and PLK1-mediated cell cycle arrest
- PMID: 35910374
- PMCID: PMC9330219
- DOI: 10.3389/fphar.2022.942261
Exogenous proline enhances susceptibility of NSCLC to cisplatin via metabolic reprogramming and PLK1-mediated cell cycle arrest
Abstract
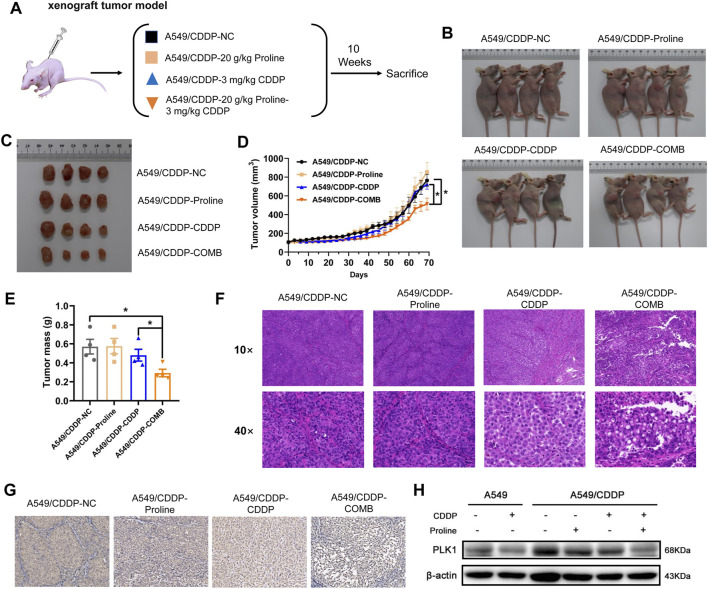
The occurrence of cisplatin resistance is still the main factor limiting the therapeutic effect of non-small cell lung cancer (NSCLC). It is urgent to elucidate the resistance mechanism and develop novel treatment strategies. Targeted metabolomics was first performed to detect amino acids' content in cisplatin-resistant cancer cells considering the relationship between tumour metabolic rearrangement and chemotherapy resistance and chemotherapy resistance. We discovered that levels of most amino acids were significantly downregulated, whereas exogenous supplementation of proline could enhance the sensitivity of NSCLC cells to cisplatin, evidenced by inhibited cell viability and tumour growth in vitro and xenograft models. In addition, the combined treatment of proline and cisplatin suppressed ATP production through disruption of the TCA cycle and oxidative phosphorylation. Furthermore, transcriptomic analysis identified the cell cycle as the top enriched pathway in co-therapy cells, accompanied by significant down-regulation of PLK1, a serine/threonine-protein kinase. Mechanistic studies revealed that PLK1 inhibitor (BI2536) and CDDP have synergistic inhibitory effects on NSCLC cells, and cells transfected with lentivirus expressing shPLK1 showed significantly increased toxicity to cisplatin. Inhibition of PLK1 inactivated AMPK, a primary regulator of cellular energy homeostasis, ultimately leading to cell cycle arrest via FOXO3A-FOXM1 axis mediated transcriptional inhibition in cisplatin-resistant cells. In conclusion, our study demonstrates that exogenous proline exerts an adjuvant therapeutic effect on cisplatin resistance, and PLK1 may be considered an attractive target for the clinical treatment of cisplatin resistance in NSCLC.
Keywords: NSCLC; cisplatin resistance; metabolic reprogramming; plk1; proline.
Copyright © 2022 Han, Sun, Zhang, Yue, Tian, Yan, Yin, Qin and Zhao.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





References
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous