

Bartonella taylorii: A Model Organism for Studying Bartonella Infection in vitro and in vivo
- PMID: 35910598
- PMCID: PMC9336547
- DOI: 10.3389/fmicb.2022.913434
Bartonella taylorii: A Model Organism for Studying Bartonella Infection in vitro and in vivo
Abstract
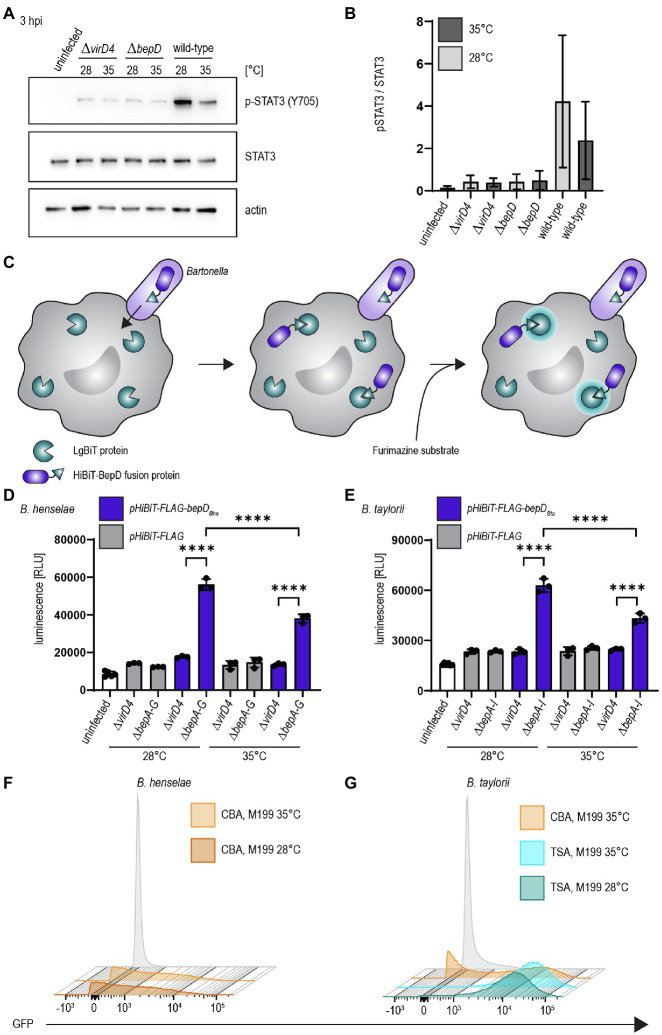
Bartonella spp. are Gram-negative facultative intracellular pathogens that infect diverse mammals and cause a long-lasting intra-erythrocytic bacteremia in their natural host. These bacteria translocate Bartonella effector proteins (Beps) into host cells via their VirB/VirD4 type 4 secretion system (T4SS) in order to subvert host cellular functions, thereby leading to the downregulation of innate immune responses. Most studies on the functional analysis of the VirB/VirD4 T4SS and the Beps were performed with the major zoonotic pathogen Bartonella henselae for which efficient in vitro infection protocols have been established. However, its natural host, the cat, is unsuitable as an experimental infection model. In vivo studies were mostly confined to rodent models using rodent-specific Bartonella species, while the in vitro infection protocols devised for B. henselae are not transferable for those pathogens. The disparities of in vitro and in vivo studies in different species have hampered progress in our understanding of Bartonella pathogenesis. Here we describe the murine-specific strain Bartonella taylorii IBS296 as a new model organism facilitating the study of bacterial pathogenesis both in vitro in cell cultures and in vivo in laboratory mice. We implemented the split NanoLuc luciferase-based translocation assay to study BepD translocation through the VirB/VirD4 T4SS. We found increased effector-translocation into host cells if the bacteria were grown on tryptic soy agar (TSA) plates and experienced a temperature shift immediately before infection. The improved infectivity in vitro was correlating to an upregulation of the VirB/VirD4 T4SS. Using our adapted infection protocols, we showed BepD-dependent immunomodulatory phenotypes in vitro. In mice, the implemented growth conditions enabled infection by a massively reduced inoculum without having an impact on the course of the intra-erythrocytic bacteremia. The established model opens new avenues to study the role of the VirB/VirD4 T4SS and the translocated Bep effectors in vitro and in vivo.
Keywords: Bartonella; NanoLuc; STAT3; effector proteins; luciferase; type IV secretion system.
Copyright © 2022 Fromm, Boegli, Ortelli, Wagner, Bohn, Malmsheimer, Wagner and Dehio.
Conflict of interest statement
AW was employed by company Maucher Jenkins, Germany. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



Similar articles
-
Translocation of YopJ family effector proteins through the VirB/VirD4 T4SS of Bartonella.Proc Natl Acad Sci U S A. 2024 May 14;121(20):e2310348121. doi: 10.1073/pnas.2310348121. Epub 2024 May 6. Proc Natl Acad Sci U S A. 2024. PMID: 38709922 Free PMC article.
-
The Impact of Bartonella VirB/VirD4 Type IV Secretion System Effectors on Eukaryotic Host Cells.Front Microbiol. 2021 Dec 15;12:762582. doi: 10.3389/fmicb.2021.762582. eCollection 2021. Front Microbiol. 2021. PMID: 34975788 Free PMC article. Review.
-
Role of distinct type-IV-secretion systems and secreted effector sets in host adaptation by pathogenic Bartonella species.Cell Microbiol. 2019 Mar;21(3):e13004. doi: 10.1111/cmi.13004. Epub 2019 Feb 6. Cell Microbiol. 2019. PMID: 30644157 Free PMC article. Review.
-
Bartonella henselae trimeric autotransporter adhesin BadA expression interferes with effector translocation by the VirB/D4 type IV secretion system.Cell Microbiol. 2013 May;15(5):759-78. doi: 10.1111/cmi.12070. Epub 2012 Dec 6. Cell Microbiol. 2013. PMID: 23163798
-
Conjugative DNA transfer into human cells by the VirB/VirD4 type IV secretion system of the bacterial pathogen Bartonella henselae.Proc Natl Acad Sci U S A. 2011 Aug 30;108(35):14643-8. doi: 10.1073/pnas.1019074108. Epub 2011 Aug 15. Proc Natl Acad Sci U S A. 2011. PMID: 21844337 Free PMC article.
Cited by
-
Experimental Infection of Ferrets with Bartonella henselae: In Search of a Novel Animal Model for Zoonotic Bartonellosis.Pathogens. 2025 Apr 26;14(5):421. doi: 10.3390/pathogens14050421. Pathogens. 2025. PMID: 40430742 Free PMC article.
-
Translocation of YopJ family effector proteins through the VirB/VirD4 T4SS of Bartonella.Proc Natl Acad Sci U S A. 2024 May 14;121(20):e2310348121. doi: 10.1073/pnas.2310348121. Epub 2024 May 6. Proc Natl Acad Sci U S A. 2024. PMID: 38709922 Free PMC article.
-
Adaptive immune defense prevents Bartonella persistence upon trans-placental transmission.PLoS Pathog. 2022 May 17;18(5):e1010489. doi: 10.1371/journal.ppat.1010489. eCollection 2022 May. PLoS Pathog. 2022. PMID: 35580143 Free PMC article.
-
Cutting Edge: Redundant Roles for MHC Class II-, CD1d-, and MR1-restricted T Cells in Clearing Bartonella Infection.J Immunol. 2024 Sep 1;213(5):553-558. doi: 10.4049/jimmunol.2400045. J Immunol. 2024. PMID: 38984869 Free PMC article.
References
-
- Boulouis H. J., Barrat F., Bermond D., Bernex F., Thibault D., Heller R., et al. . (2001). Kinetics of Bartonella birtlesii infection in experimentally infected mice and pathogenic effect on reproductive functions. Infect. Immun. 69, 5313–5317. doi: 10.1128/iai.69.9.5313-5317.2001, PMID: - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous