

Dedifferentiated Schwann cell-derived TGF-β3 is essential for the neural system to promote wound healing
- PMID: 35910794
- PMCID: PMC9330527
- DOI: 10.7150/thno.72317
Dedifferentiated Schwann cell-derived TGF-β3 is essential for the neural system to promote wound healing
Abstract
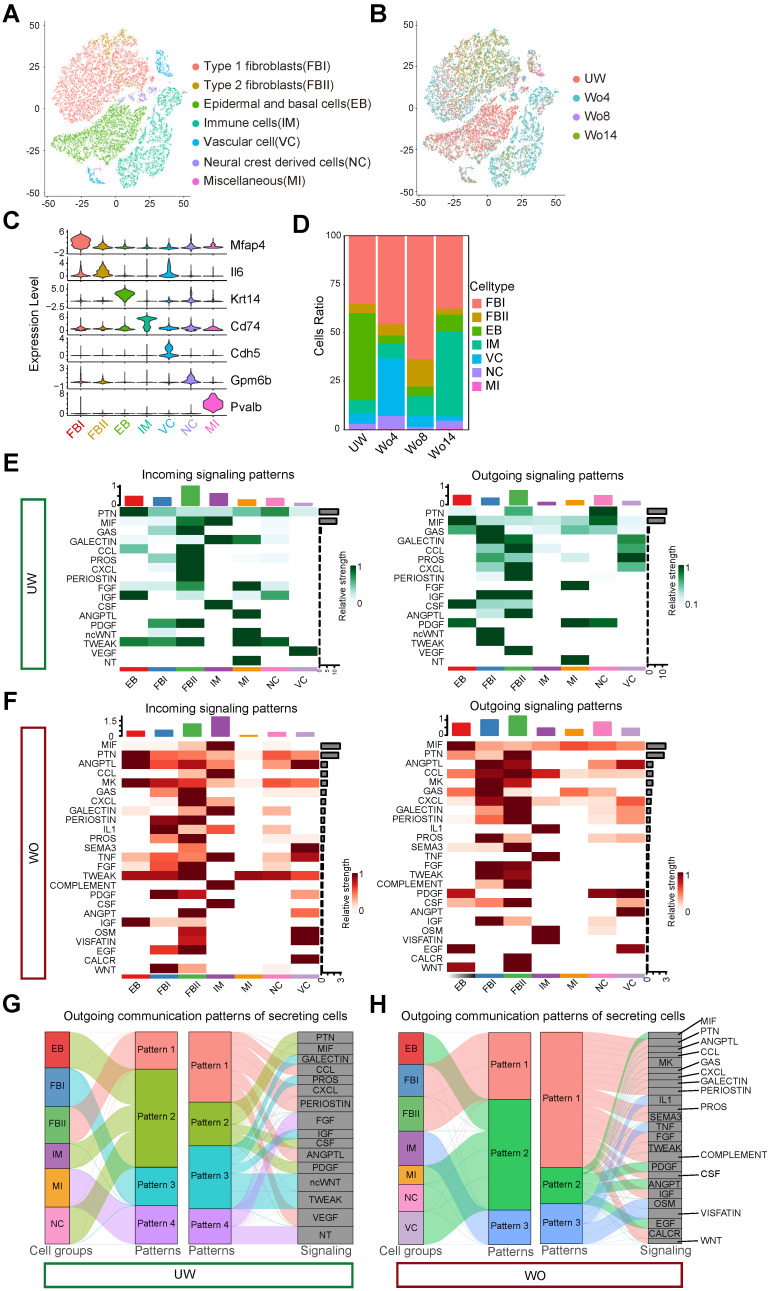
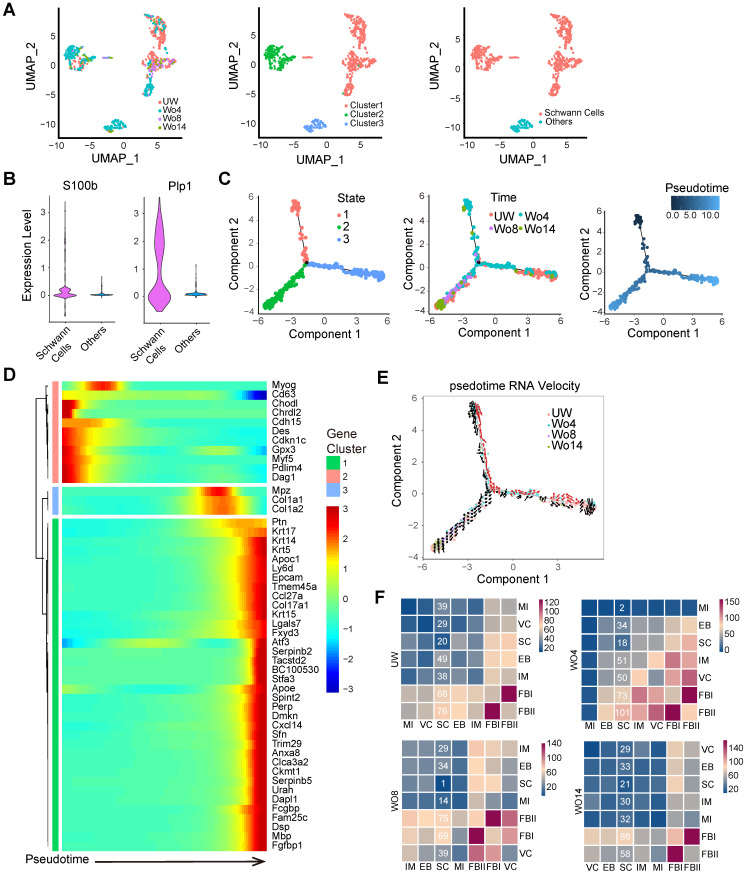
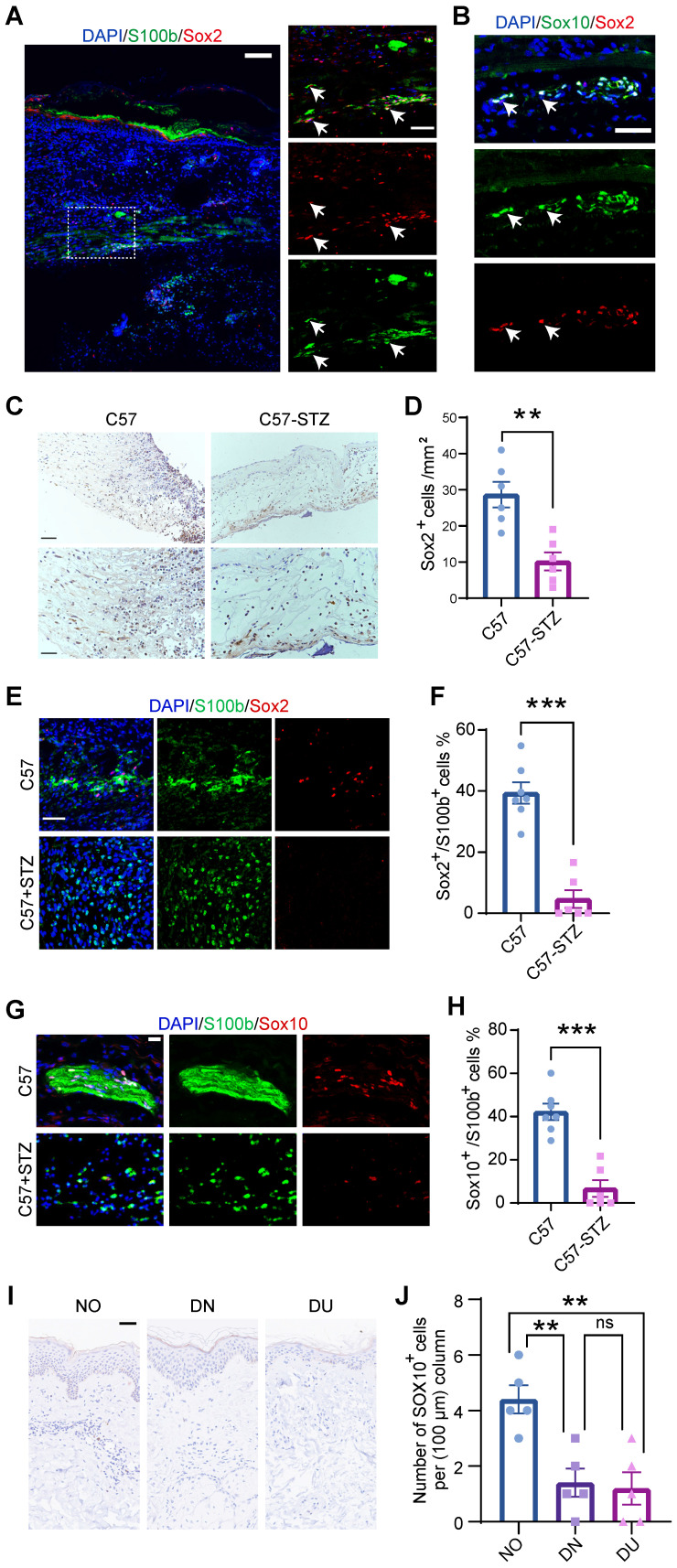
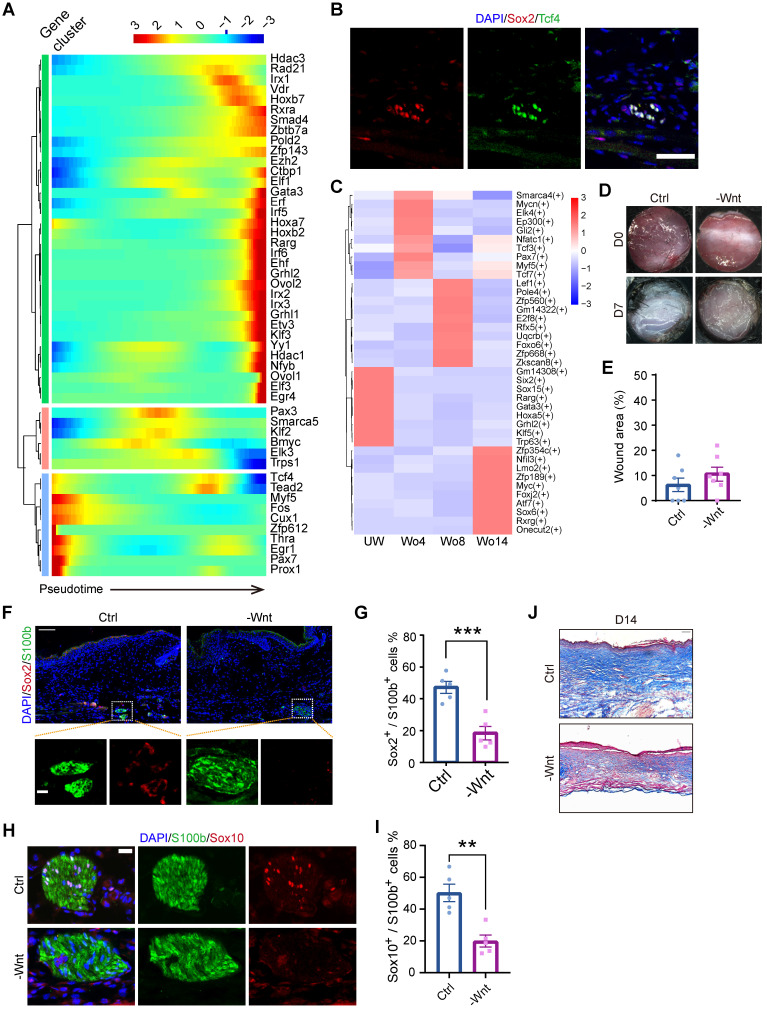
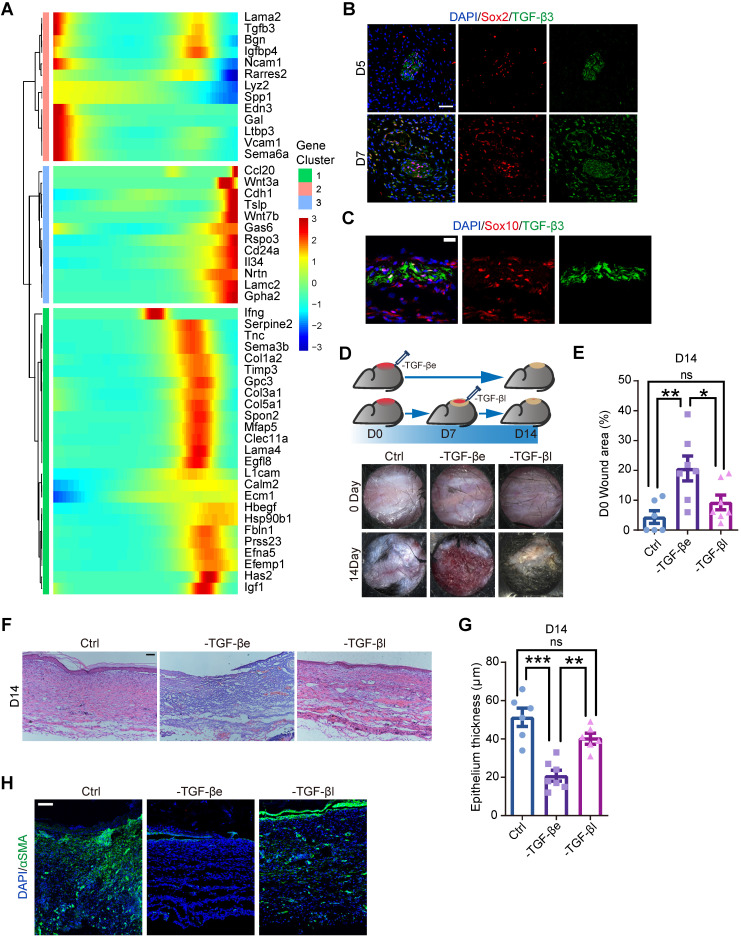
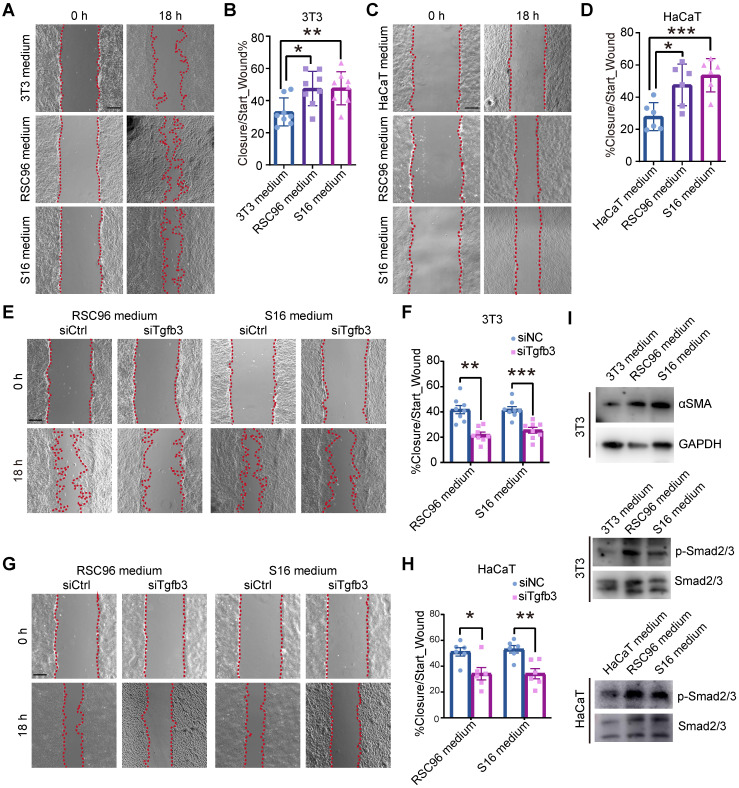
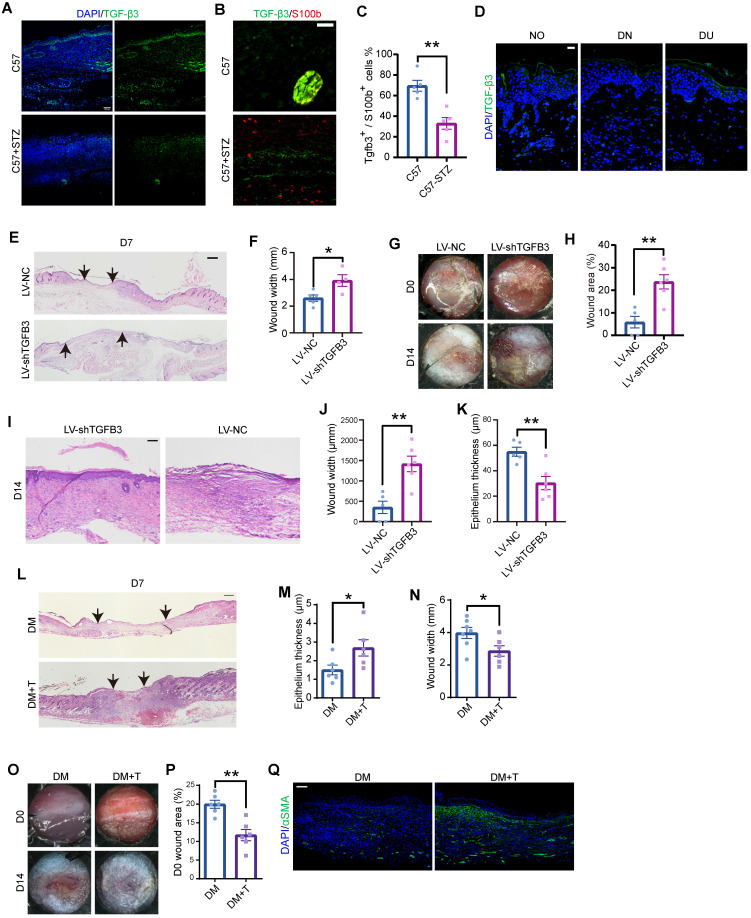
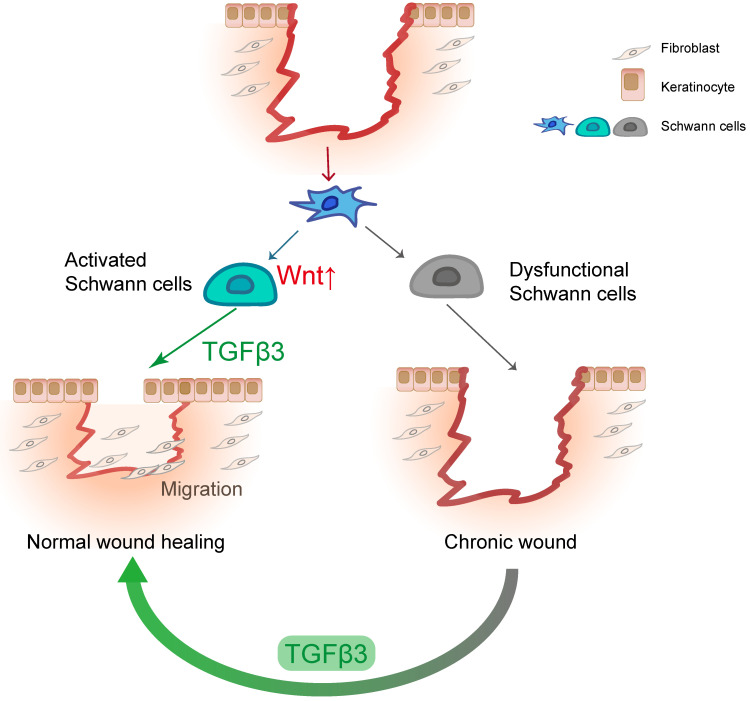
Rationale: Wound healing is among the most complicated physiological processes and requires the synchronization of various cell types with distinct roles to re-establish the condition of the original skin. Patients affected by peripheral neuropathies often experience failure to heal. Loss of Schwann cells (SCs), a crucial population of peripheral nervous system cells in skin, may contribute to chronic wounds. However, the role of SCs in wound healing are poorly understood. Methods: The activity of SCs was investigated by using a cell atlas of the wound healing process, which was generated by integrating single-cell RNA sequencing (scRNA-seq) libraries covering different states of mouse back skin. The results of in silico analysis were validated by in vitro cell culture and in vivo mouse model. Selective inhibitors and conditional RNAi by virus transfection were utilized to investigate the role of SCs in wound healing. Findings from mouse experiments were further verified in scRNA-seq analysis of diabetic patients. Results: Our in silico analysis revealed the heterogeneous cellular components of skin and the dynamic interactions of neural crest derived cells (NCs) with other cell types. We found that SCs dedifferentiated at an early stage of wound repair with upregulated Wnt signaling. We also identified dedifferentiated SC (dSC) defect in diabetic wounds in both mouse and human. Wnt inhibition at the wound site repressed SC dedifferentiation, leading to defective repair. Furthermore, dSCs derived TGF-β3, which is context-dependent, promoted the migration of fibroblasts and keratinocytes. Moreover, TGF-β3 supplementation enhanced the healing of chronic wounds in diabetic mice with impaired SCs. Conclusion: Our study thus advances the understanding of the roles of neural-derived cells in skin regeneration and suggests a potential therapeutic strategy for wound healing disorders.
Keywords: Schwann cells; Single-cell transcriptome; TGF-β; skin; wound repair.
© The author(s).
Conflict of interest statement
Competing Interests: The authors have declared that no competing interest exists.
Figures
Similar articles
-
A Dermal Equivalent Engineered with TGF-β3 Expressing Bone Marrow Stromal Cells and Amniotic Membrane: Cosmetic Healing of Full-Thickness Skin Wounds in Rats.Artif Organs. 2016 Dec;40(12):E266-E279. doi: 10.1111/aor.12807. Artif Organs. 2016. PMID: 27911032
-
Decoding the complexity of delayed wound healing following Enterococcus faecalis infection.Elife. 2024 May 20;13:RP95113. doi: 10.7554/eLife.95113. Elife. 2024. PMID: 38767331 Free PMC article.
-
Diminished schwann cell repair responses play a role in delayed diabetes-associated wound healing.Front Physiol. 2022 Dec 22;13:814754. doi: 10.3389/fphys.2022.814754. eCollection 2022. Front Physiol. 2022. PMID: 36620211 Free PMC article.
-
Scar-free healing: from embryonic mechanisms to adult therapeutic intervention.Philos Trans R Soc Lond B Biol Sci. 2004 May 29;359(1445):839-50. doi: 10.1098/rstb.2004.1475. Philos Trans R Soc Lond B Biol Sci. 2004. PMID: 15293811 Free PMC article. Review.
-
[The modern approach to wound treatment].Med Pregl. 2000 Jul-Aug;53(7-8):363-8. Med Pregl. 2000. PMID: 11214479 Review. Croatian.
Cited by
-
Characterizing Fibroblast Heterogeneity in Diabetic Wounds Through Single-Cell RNA-Sequencing.Biomedicines. 2024 Nov 7;12(11):2538. doi: 10.3390/biomedicines12112538. Biomedicines. 2024. PMID: 39595104 Free PMC article. Review.
-
Alteration of skin fibroblast steady state contributes to healing outcomes.bioRxiv [Preprint]. 2024 Dec 12:2024.12.06.627278. doi: 10.1101/2024.12.06.627278. bioRxiv. 2024. PMID: 39713414 Free PMC article. Preprint.
-
Cell Heterogeneity and Variability in Peripheral Nerve after Injury.Int J Mol Sci. 2024 Mar 20;25(6):3511. doi: 10.3390/ijms25063511. Int J Mol Sci. 2024. PMID: 38542483 Free PMC article. Review.
-
Marine bioactive peptides as potential therapeutic agents for wound healing - a review.Ann Med. 2025 Dec;57(1):2530693. doi: 10.1080/07853890.2025.2530693. Epub 2025 Jul 13. Ann Med. 2025. PMID: 40652403 Free PMC article. Review.
-
Human Parathyroid Hormone Analog (3-34/29-34) promotes wound re-epithelialization through inducing keratinocyte migration and epithelial-mesenchymal transition via PTHR1-PI3K/AKT activation.Cell Commun Signal. 2023 Aug 23;21(1):217. doi: 10.1186/s12964-023-01243-9. Cell Commun Signal. 2023. PMID: 37612710 Free PMC article.
References
-
- Carr MJ, Toma JS, Johnston APW, Steadman PE, Yuzwa SA, Mahmud N. et al. Mesenchymal precursor cells in adult nerves contribute to mammalian tissue repair and regeneration. Cell Stem Cell. 2019;24:240–256. e249. - PubMed
-
- Barker AR, Rosson GD, Dellon AL. Wound healing in denervated tissue. Ann Plast Surg. 2006;57:339–342. - PubMed
-
- Harsum S, Clarke JD, Martin P. A reciprocal relationship between cutaneous nerves and repairing skin wounds in the developing chick embryo. Dev Biol. 2001;238:27–39. - PubMed
-
- Chéret J, Lebonvallet N, Buhé V, Carre JL, Misery L, Le Gall-Ianotto C. Influence of sensory neuropeptides on human cutaneous wound healing process. J Dermatol Sci. 2014;74:193–203. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
