

Moonlighting at the Poles: Non-Canonical Functions of Centrosomes
- PMID: 35912107
- PMCID: PMC9329689
- DOI: 10.3389/fcell.2022.930355
Moonlighting at the Poles: Non-Canonical Functions of Centrosomes
Abstract
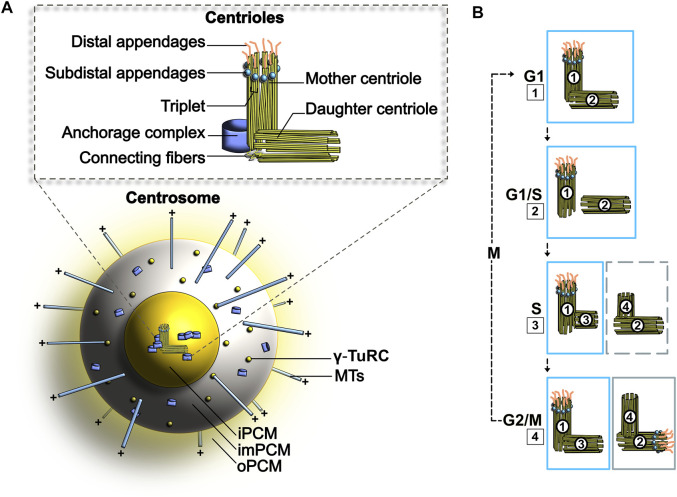
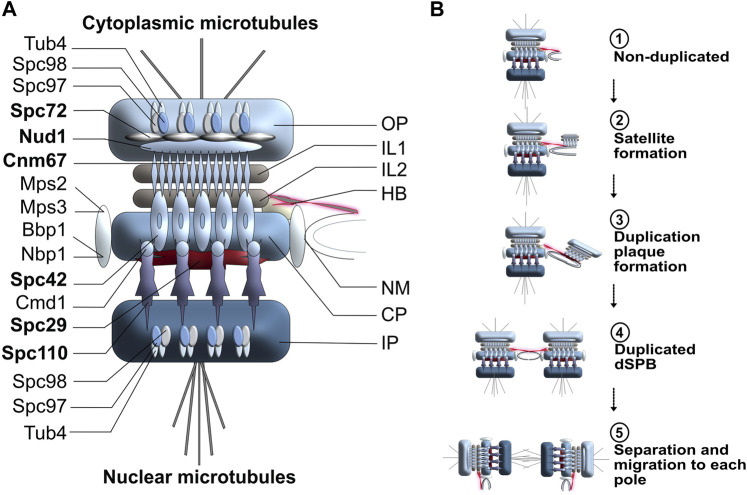
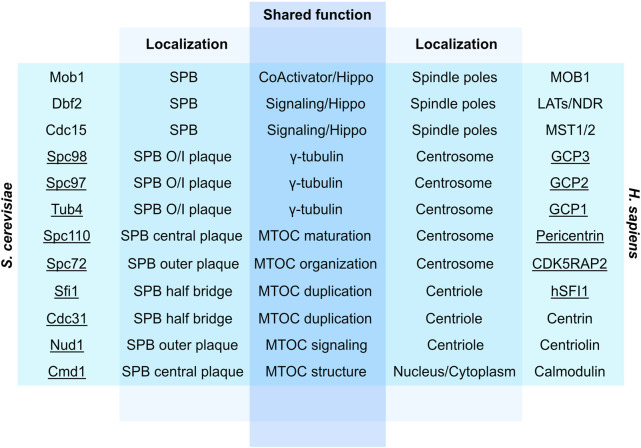
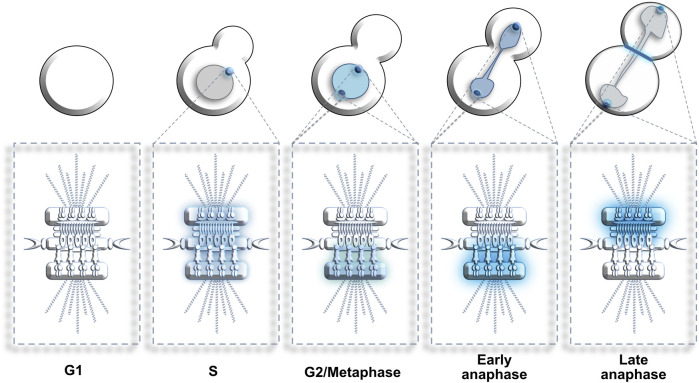
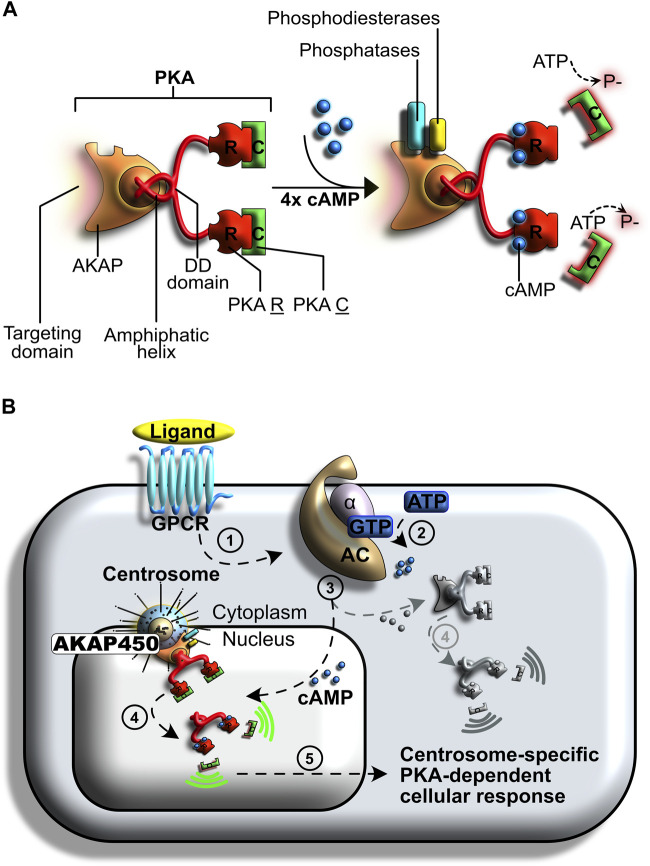
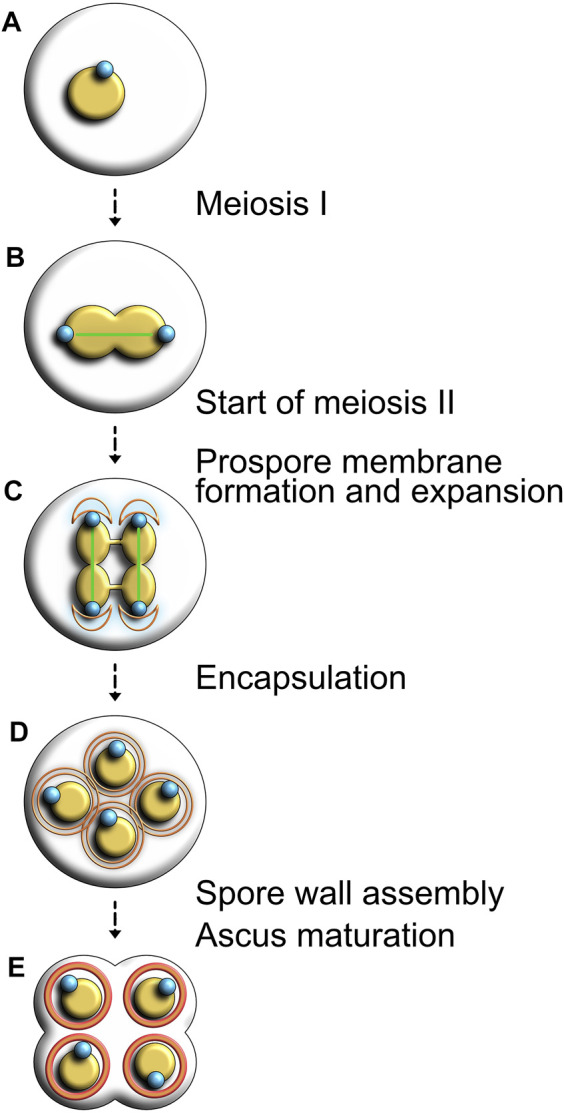
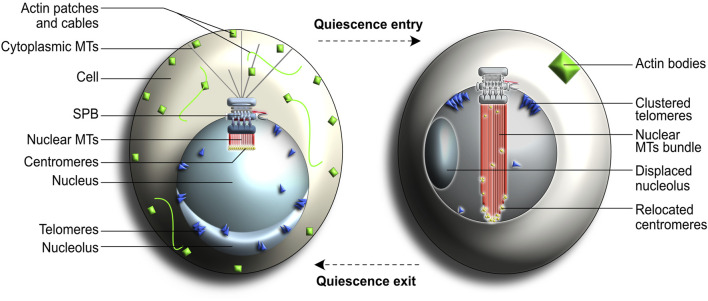
Centrosomes are best known as the microtubule organizing centers (MTOCs) of eukaryotic cells. In addition to their classic role in chromosome segregation, centrosomes play diverse roles unrelated to their MTOC activity during cell proliferation and quiescence. Metazoan centrosomes and their functional doppelgängers from lower eukaryotes, the spindle pole bodies (SPBs), act as important structural platforms that orchestrate signaling events essential for cell cycle progression, cellular responses to DNA damage, sensory reception and cell homeostasis. Here, we provide a critical overview of the unconventional and often overlooked roles of centrosomes/SPBs in the life cycle of eukaryotic cells.
Keywords: Cdc5; MTOCs; PLK1; cell cycle; centrosomes; spindle pole bodies.
Copyright © 2022 Langlois-Lemay and D’Amours.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Yeast centrosomes act as organizing centers to promote Polo kinase-mediated adaptation to persistent DNA damage.Proc Natl Acad Sci U S A. 2025 Jul 8;122(27):e2414426122. doi: 10.1073/pnas.2414426122. Epub 2025 Jul 2. Proc Natl Acad Sci U S A. 2025. PMID: 40601630
-
CIP2A acts as a scaffold for CEP192-mediated microtubule organizing center assembly by recruiting Plk1 and aurora A during meiotic maturation.Development. 2017 Oct 15;144(20):3829-3839. doi: 10.1242/dev.158584. Epub 2017 Sep 21. Development. 2017. PMID: 28935709
-
Application of PALM Superresolution Microscopy to the Analysis of Microtubule-Organizing Centers (MTOCs) in Aspergillus nidulans.Methods Mol Biol. 2021;2329:277-289. doi: 10.1007/978-1-0716-1538-6_20. Methods Mol Biol. 2021. PMID: 34085230
-
Control at the cell center: the role of spindle poles in cytoskeletal organization and cell cycle regulation.Cell Cycle. 2007 Nov 15;6(22):2788-94. doi: 10.4161/cc.6.22.4941. Epub 2007 Aug 27. Cell Cycle. 2007. PMID: 17986861 Review.
-
A guiding torch at the poles: the multiple roles of spindle microtubule-organizing centers during cell division.Cell Cycle. 2020 Jun;19(12):1405-1421. doi: 10.1080/15384101.2020.1754586. Epub 2020 May 13. Cell Cycle. 2020. PMID: 32401610 Free PMC article. Review.
Cited by
-
Amplified centrosomes-more than just a threat.EMBO Rep. 2024 Oct;25(10):4153-4167. doi: 10.1038/s44319-024-00260-0. Epub 2024 Sep 16. EMBO Rep. 2024. PMID: 39285247 Free PMC article. Review.
-
Yeast centrosomes act as organizing centers to promote Polo kinase-mediated adaptation to persistent DNA damage.Proc Natl Acad Sci U S A. 2025 Jul 8;122(27):e2414426122. doi: 10.1073/pnas.2414426122. Epub 2025 Jul 2. Proc Natl Acad Sci U S A. 2025. PMID: 40601630
-
New mutations in the core Schizosaccharomyces pombe spindle pole body scaffold Ppc89 reveal separable functions in regulating cell division.G3 (Bethesda). 2025 Jan 8;15(1):jkae249. doi: 10.1093/g3journal/jkae249. G3 (Bethesda). 2025. PMID: 39471327 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous