

Revealing a Hidden Intermediate of Rotatory Catalysis with X-ray Crystallography and Molecular Simulations
- PMID: 35912346
- PMCID: PMC9336149
- DOI: 10.1021/acscentsci.1c01599
Revealing a Hidden Intermediate of Rotatory Catalysis with X-ray Crystallography and Molecular Simulations
Abstract
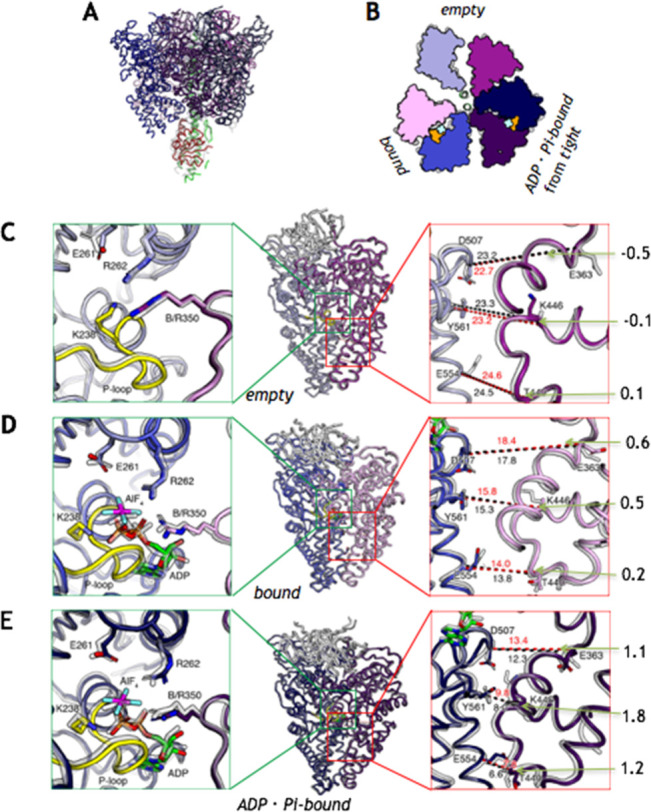
The mechanism of rotatory catalysis in ATP-hydrolyzing molecular motors remains an unresolved puzzle in biological energy transfer. Notwithstanding the wealth of available biochemical and structural information inferred from years of experiments, knowledge on how the coupling between the chemical and mechanical steps within motors enforces directional rotatory movements remains fragmentary. Even more contentious is to pinpoint the rate-limiting step of a multistep rotation process. Here, using vacuolar or V1-type hexameric ATPase as an exemplary rotational motor, we present a model of the complete 4-step conformational cycle involved in rotatory catalysis. First, using X-ray crystallography, a new intermediate or "dwell" is identified, which enables the release of an inorganic phosphate (or Pi) after ATP hydrolysis. Using molecular dynamics simulations, this new dwell is placed in a sequence with three other crystal structures to derive a putative cyclic rotation path. Free-energy simulations are employed to estimate the rate of the hexameric protein transformations and delineate allosteric effects that allow new reactant ATP entry only after hydrolysis product exit. An analysis of transfer entropy brings to light how the side-chain-level interactions transcend into larger-scale reorganizations, highlighting the role of the ubiquitous arginine-finger residues in coupling chemical and mechanical information. An inspection of all known rates encompassing the 4-step rotation mechanism implicates the overcoming of the ADP interactions with V1-ATPase to be the rate-limiting step of motor action.
© 2022 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures






Similar articles
-
Chemomechanical Coupling in Hexameric Protein-Protein Interfaces Harnesses Energy within V-Type ATPases.J Am Chem Soc. 2017 Jan 11;139(1):293-310. doi: 10.1021/jacs.6b10744. Epub 2016 Dec 23. J Am Chem Soc. 2017. PMID: 27936329 Free PMC article.
-
Rotational Mechanism Model of the Bacterial V1 Motor Based on Structural and Computational Analyses.Front Physiol. 2019 Feb 5;10:46. doi: 10.3389/fphys.2019.00046. eCollection 2019. Front Physiol. 2019. PMID: 30804798 Free PMC article. Review.
-
Mechanism of substrate translocation by a ring-shaped ATPase motor at millisecond resolution.J Am Chem Soc. 2015 Mar 4;137(8):3031-40. doi: 10.1021/ja512605w. Epub 2015 Feb 19. J Am Chem Soc. 2015. PMID: 25646698 Free PMC article.
-
Single-molecule analysis reveals rotational substeps and chemo-mechanical coupling scheme of Enterococcus hirae V1-ATPase.J Biol Chem. 2019 Nov 8;294(45):17017-17030. doi: 10.1074/jbc.RA119.008947. Epub 2019 Sep 13. J Biol Chem. 2019. PMID: 31519751 Free PMC article.
-
Structure and dynamics of rotary V1 motor.Cell Mol Life Sci. 2018 May;75(10):1789-1802. doi: 10.1007/s00018-018-2758-3. Epub 2018 Jan 31. Cell Mol Life Sci. 2018. PMID: 29387903 Free PMC article. Review.
Cited by
-
Physical pictures of rotation mechanisms of F1- and V1-ATPases: Leading roles of translational, configurational entropy of water.Front Mol Biosci. 2023 Jun 9;10:1159603. doi: 10.3389/fmolb.2023.1159603. eCollection 2023. Front Mol Biosci. 2023. PMID: 37363397 Free PMC article.
-
Six states of Enterococcus hirae V-type ATPase reveals non-uniform rotor rotation during turnover.Commun Biol. 2023 Jul 28;6(1):755. doi: 10.1038/s42003-023-05110-8. Commun Biol. 2023. PMID: 37507515 Free PMC article.
-
Na+-V-ATPase inhibitor curbs VRE growth and unveils Na+ pathway structure.Nat Struct Mol Biol. 2025 Mar;32(3):450-458. doi: 10.1038/s41594-024-01419-y. Epub 2024 Nov 21. Nat Struct Mol Biol. 2025. PMID: 39572733
-
ATP synthesis of Enterococcus hirae V-ATPase driven by sodium motive force.J Biol Chem. 2025 Apr;301(4):108422. doi: 10.1016/j.jbc.2025.108422. Epub 2025 Mar 19. J Biol Chem. 2025. PMID: 40118453 Free PMC article.
References
-
- Arai S.; Saijo S.; Suzuki K.; Mizutani K.; Kakinuma Y.; Ishizuka-Katsura Y.; Ohsawa N.; Terada T.; Shirouzu M.; Yokoyama S.; Iwata S.; Yamato I.; Murata T. Rotation Mechanism of Enterococcus Hirae V1-ATPase Based on Asymmetric Crystal Structures. Nature 2013, 493 (7434), 703–707. 10.1038/nature11778. - DOI - PubMed
-
- Suzuki K.; Mizutani K.; Maruyama S.; Shimono K.; Imai F. L.; Muneyuki E.; Kakinuma Y.; Ishizuka-Katsura Y.; Shirouzu M.; Yokoyama S.; Yamato I.; Murata T. Crystal Structures of the ATP-Binding and ADP-Release Dwells of the V1 Rotary Motor. Nat. Commun. 2016, 7, 13235.10.1038/ncomms13235. - DOI - PMC - PubMed
-
- Maruyama S.; Suzuki K.; Imamura M.; Sasaki H.; Matsunami H.; Mizutani K.; Saito Y.; Imai F. L.; Ishizuka-Katsura Y.; Kimura-Someya T.; Shirouzu M.; Uchihashi T.; Ando T.; Yamato I.; Murata T. Metastable Asymmetrical Structure of a Shaftless V1Motor. Sci. Adv. 2019, 5 (1), eaau814910.1126/sciadv.aau8149. - DOI - PMC - PubMed
-
- Saijo S.; Arai S.; Hossain K. M. M.; Yamato I.; Suzuki K.; Kakinuma Y.; Ishizuka-Katsura Y.; Ohsawa N.; Terada T.; Shirouzu M.; Yokoyama S.; Iwata S.; Murata T. Crystal Structure of the Central Axis DF Complex of the Prokaryotic V-ATPase. Proc. Natl. Acad. Sci. U. S. A. 2011, 108 (50), 19955–19960. 10.1073/pnas.1108810108. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
