

Structure and Function of BcpE2, the Most Promiscuous GH3-Family Glucose Scavenging Beta-Glucosidase
- PMID: 35913158
- PMCID: PMC9426481
- DOI: 10.1128/mbio.00935-22
Structure and Function of BcpE2, the Most Promiscuous GH3-Family Glucose Scavenging Beta-Glucosidase
Abstract
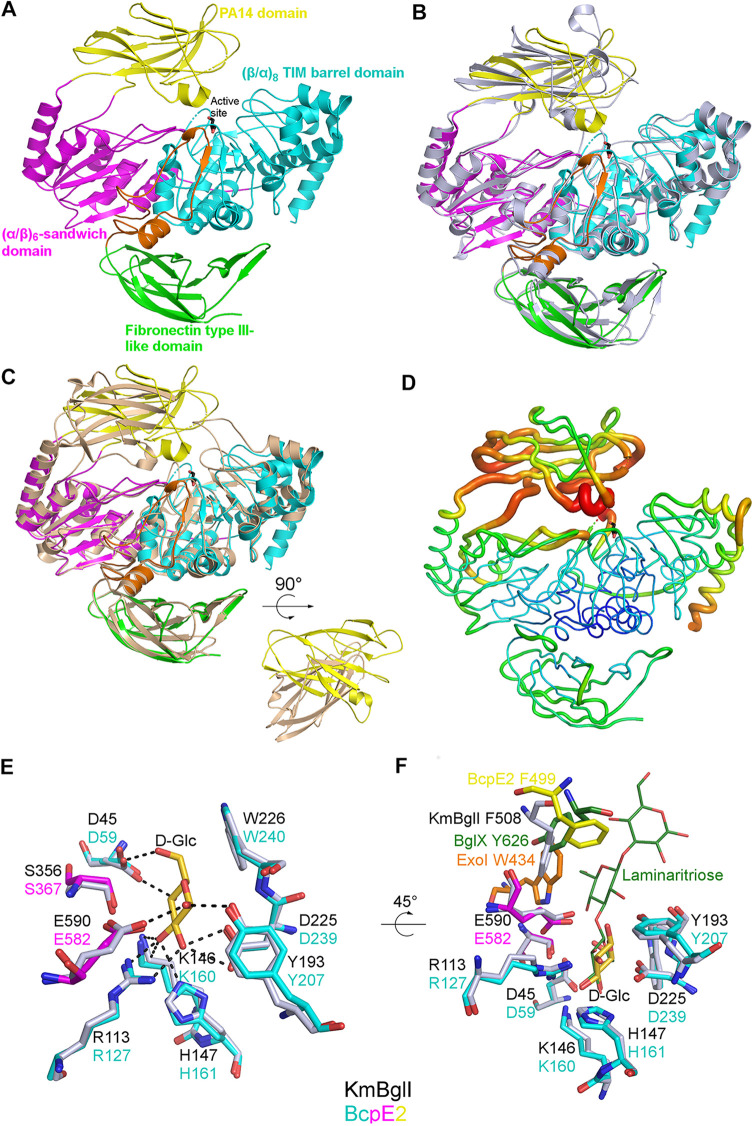
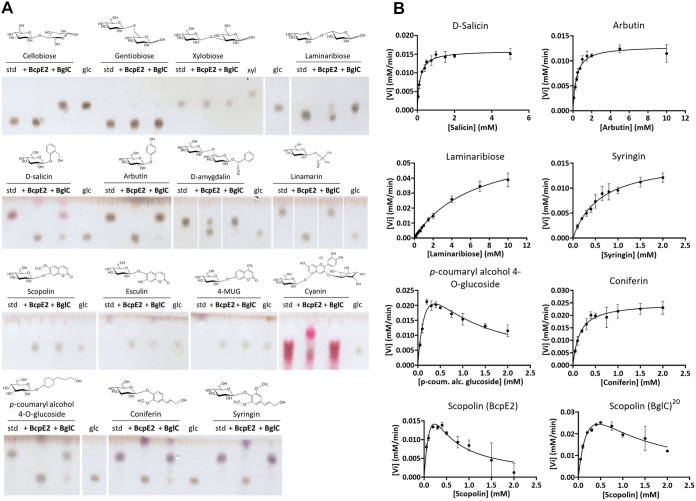
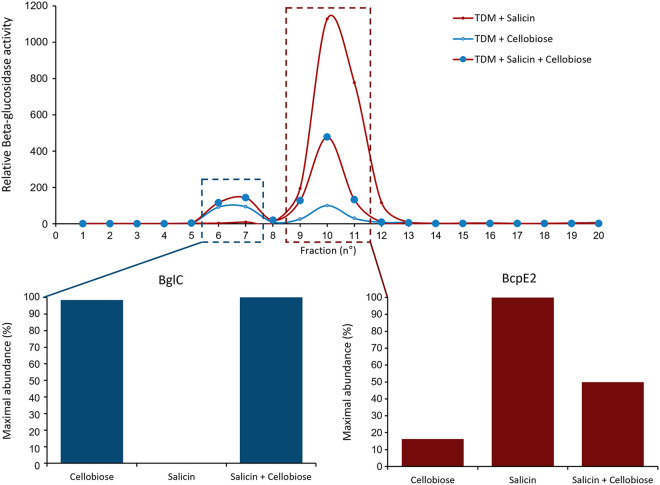
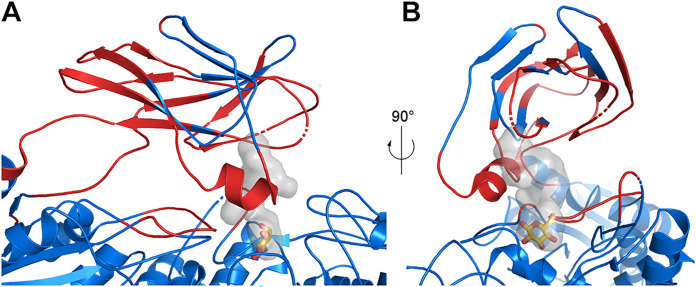
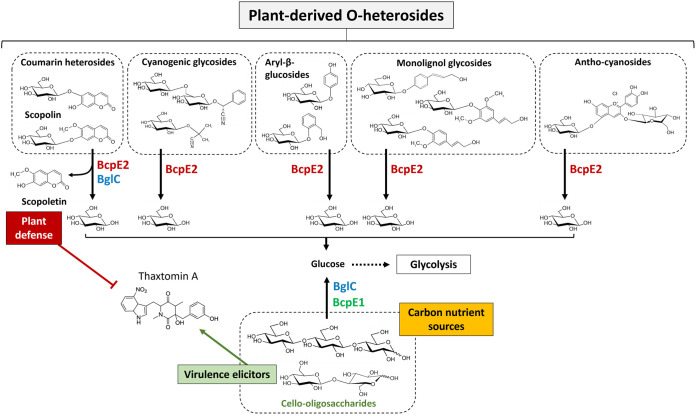
Cellulose being the most abundant polysaccharide on earth, beta-glucosidases hydrolyzing cello-oligosaccharides are key enzymes to fuel glycolysis in microorganisms developing on plant material. In Streptomyces scabiei, the causative agent of common scab in root and tuber crops, a genetic compensation phenomenon safeguards the loss of the gene encoding the cello-oligosaccharide hydrolase BglC by awakening the expression of alternative beta-glucosidases. Here, we revealed that the BglC compensating enzyme BcpE2 was the GH3-family beta-glucosidase that displayed the highest reported substrate promiscuity and was able to release the glucose moiety of all tested types of plant-derived heterosides (aryl β-glucosides, monolignol glucosides, cyanogenic glucosides, anthocyanosides, and coumarin heterosides). BcpE2 structure analysis highlighted a large cavity in the PA14 domain that covered the active site, and the high flexibility of this domain would allow proper adjustment of this cavity for disparate heterosides. The exceptional substrate promiscuity of BcpE2 provides microorganisms a versatile tool for scavenging glucose from plant-derived nutrients that widely vary in size and structure. Importantly, scopolin was the only substrate commonly hydrolyzed by both BglC and BcpE2, thereby generating the potent virulence inhibitor scopoletin. Next to fueling glycolysis, both enzymes would also fine-tune the strength of virulence. IMPORTANCE Plant decaying biomass is the most abundant provider of carbon sources for soil-dwelling microorganisms. To optimally evolve in such environmental niches, microorganisms possess an arsenal of hydrolytic enzymatic complexes to feed on the various types of polysaccharides, oligosaccharides, and monosaccharides. In this work, structural, enzymatic, and expression studies revealed the existence of a "swiss-army knife" enzyme, BcpE2, that was able to retrieve the glucose moiety of a multitude of plant-derived substrates that vary in size, structure, and origin. This enzyme would provide the microorganisms with a tool that would allow them to find nutrients from any type of plant-derived material.
Keywords: carbon metabolism; enzyme promiscuity; genetic compensation; glycosyl hydrolase; host-pathogen interaction; plant heterosides.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Book AJ, Lewin GR, McDonald BR, Takasuka TE, Wendt-Pienkowski E, Doering DT, Suh S, Raffa KF, Fox BG, Currie CR. 2016. Evolution of high cellulolytic activity in symbiotic streptomyces through selection of expanded gene content and coordinated gene expression. PLoS Biol 14:e1002475. doi:10.1371/journal.pbio.1002475. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
