

Impact of SARS-CoV-2 Spike Mutations on Its Activation by TMPRSS2 and the Alternative TMPRSS13 Protease
- PMID: 35913162
- PMCID: PMC9426466
- DOI: 10.1128/mbio.01376-22
Impact of SARS-CoV-2 Spike Mutations on Its Activation by TMPRSS2 and the Alternative TMPRSS13 Protease
Abstract
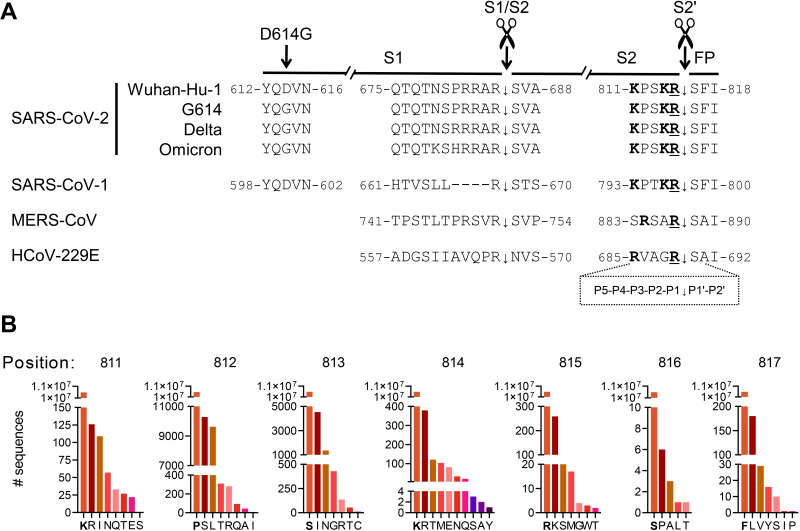
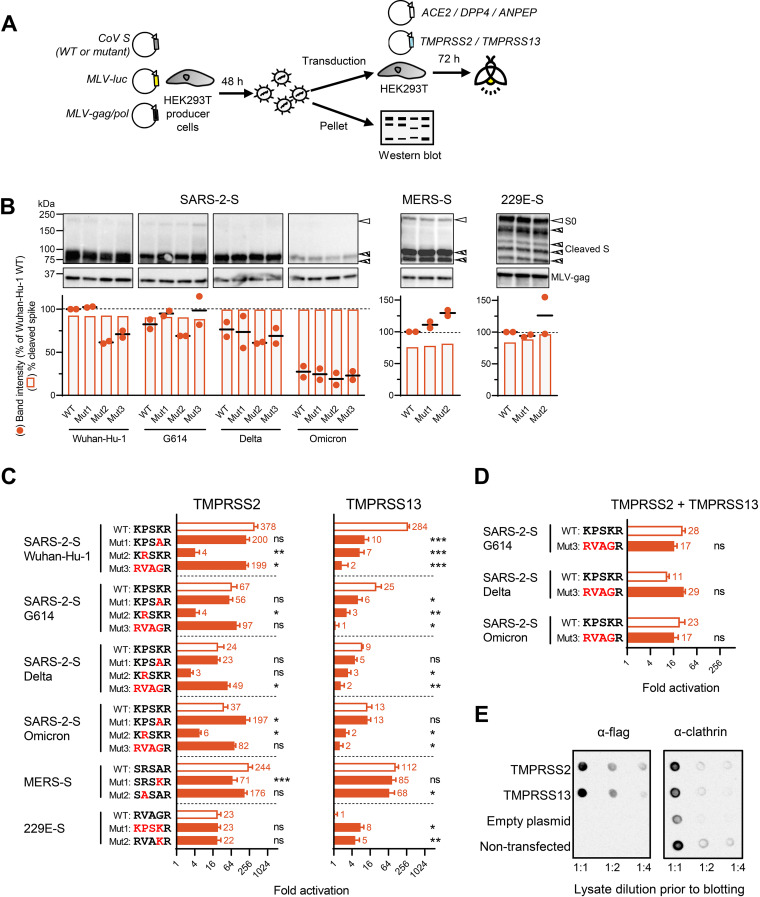
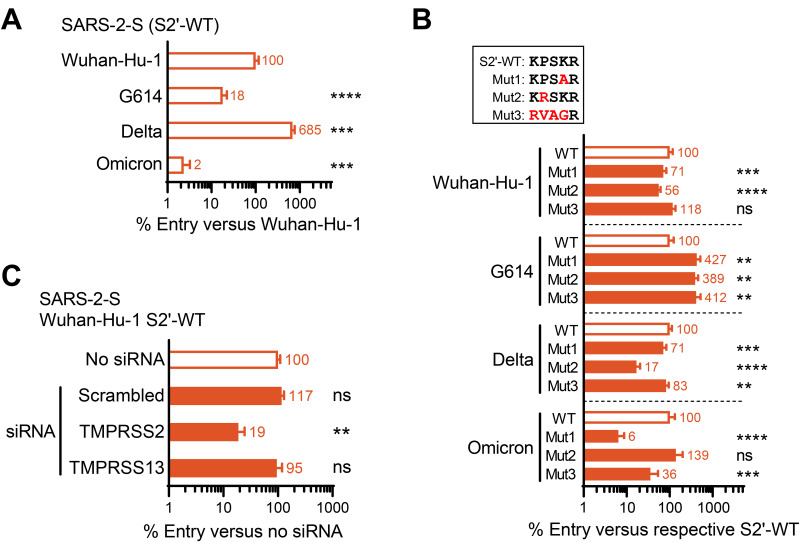
The continuous emergence of new variants of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) urges better understanding of the functional motifs in the spike (S) protein and their tolerance to mutations. Here, we focused on the S2' motif, which, during virus entry, requires cleavage by a host cell protease to release the fusion peptide. Though belonging to an immunogenic region, the SARS-CoV-2 S2' motif (811-KPSKR-815) has shown hardly any variation, with its three basic (K/R) residues being >99.99% conserved thus far. By creating a series of mutant pseudoviruses bearing the spikes of Wuhan-Hu-1, its G614 mutant or the Delta and Omicron variants, we show that residue K814 (preceding the scissile R815) is dispensable for TMPRSS2 yet favored by the alternative TMPRSS13 protease. Activation by TMPRSS13 was drastically reduced when the SARS-CoV-2 S2' motif was swapped with that of the low pathogenic 229E coronavirus (685-RVAGR-689), and also, the reverse effect was seen. This swap had no impact on recognition by TMPRSS2. In the Middle East respiratory syndrome coronavirus (MERS-CoV) spike, introducing a dibasic scissile motif was easily accepted by TMPRSS13 but less so by TMPRSS2, confirming that TMPRSS13 favors a sequence rich in K/R residues. Pseudovirus entry experiments in Calu-3 cells confirmed that the S2' mutations have minor impact on TMPRSS2. Our findings are the first to demonstrate which S2' residues are important for SARS-CoV-2 spike activation by these two airway proteases, with TMPRSS2 being more tolerant to variation than TMPRSS13. This preemptive insight will help to estimate the impact of S2' motif changes as they appear in new SARS-CoV-2 variants. IMPORTANCE Since its introduction in humans, SARS-CoV-2 is evolving with frequent appearance of new variants. The surveillance would benefit from proactive characterization of the functional motifs in the spike (S) protein, the most variable viral factor. This is linked to immune evasion but also influences spike functioning. Remarkably, though located in a strongly immunogenic region, the S2' cleavage motif has, thus far, remained highly conserved. This suggests that its sequence is critical for spike activation by airway proteases. To investigate this, we assessed how pseudovirus entry is affected by changes in the S2' motif. We demonstrate that TMPRSS2 readily accepts variations in this motif, whereas the alternative TMPRSS13 protease is more fastidious. The Wuhan-Hu-1, G614, Delta and Omicron spikes showed no difference in this regard. Being the first in its kind, our study will help to assess the impact of S2' variations as soon as they are detected during variant surveillance.
Keywords: SARS-CoV-2; TMPRSS13; TMPRSS2; cleavage; human coronavirus 229E; mutation; protease; spike protein.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The SARS-CoV-2 and other human coronavirus spike proteins are fine-tuned towards temperature and proteases of the human airways.PLoS Pathog. 2021 Apr 22;17(4):e1009500. doi: 10.1371/journal.ppat.1009500. eCollection 2021 Apr. PLoS Pathog. 2021. PMID: 33886690 Free PMC article.
-
Distinctive Roles of Furin and TMPRSS2 in SARS-CoV-2 Infectivity.J Virol. 2022 Apr 27;96(8):e0012822. doi: 10.1128/jvi.00128-22. Epub 2022 Mar 28. J Virol. 2022. PMID: 35343766 Free PMC article.
-
SARS-CoV-2 Spike Furin Cleavage Site and S2' Basic Residues Modulate the Entry Process in a Host Cell-Dependent Manner.J Virol. 2022 Jul 13;96(13):e0047422. doi: 10.1128/jvi.00474-22. Epub 2022 Jun 9. J Virol. 2022. PMID: 35678602 Free PMC article.
-
Proteolytic activation of SARS-CoV-2 spike protein.Microbiol Immunol. 2022 Jan;66(1):15-23. doi: 10.1111/1348-0421.12945. Epub 2021 Oct 12. Microbiol Immunol. 2022. PMID: 34561887 Free PMC article. Review.
-
Molecular mechanisms for understanding the association between TMPRSS2 and beta coronaviruses SARS-CoV-2, SARS-CoV and MERS-CoV infection: scoping review.Arch Microbiol. 2021 Dec 25;204(1):77. doi: 10.1007/s00203-021-02727-3. Arch Microbiol. 2021. PMID: 34953136 Free PMC article.
Cited by
-
Soluble SARS-CoV-2 Spike glycoprotein: considering some potential pathogenic effects.Front Immunol. 2025 Jun 4;16:1616106. doi: 10.3389/fimmu.2025.1616106. eCollection 2025. Front Immunol. 2025. PMID: 40534870 Free PMC article. Review.
-
Human parainfluenza virus 3 field strains undergo extracellular fusion protein cleavage to activate entry.mBio. 2024 Nov 13;15(11):e0232724. doi: 10.1128/mbio.02327-24. Epub 2024 Oct 9. mBio. 2024. PMID: 39382296 Free PMC article.
-
Functional and antigenic characterization of SARS-CoV-2 spike fusion peptide by deep mutational scanning.Nat Commun. 2024 May 14;15(1):4056. doi: 10.1038/s41467-024-48104-8. Nat Commun. 2024. PMID: 38744813 Free PMC article.
-
Functional and antigenic characterization of SARS-CoV-2 spike fusion peptide by deep mutational scanning.bioRxiv [Preprint]. 2023 Nov 29:2023.11.28.569051. doi: 10.1101/2023.11.28.569051. bioRxiv. 2023. Update in: Nat Commun. 2024 May 14;15(1):4056. doi: 10.1038/s41467-024-48104-8. PMID: 38076875 Free PMC article. Updated. Preprint.
-
Development of ketobenzothiazole-based peptidomimetic TMPRSS13 inhibitors with low nanomolar potency.J Enzyme Inhib Med Chem. 2025 Dec;40(1):2466841. doi: 10.1080/14756366.2025.2466841. Epub 2025 Feb 20. J Enzyme Inhib Med Chem. 2025. PMID: 39976239 Free PMC article.
References
-
- Zhang L, Jackson CB, Mou H, Ojha A, Peng H, Quinlan BD, Rangarajan ES, Pan A, Vanderheiden A, Suthar MS, Li W, Izard T, Rader C, Farzan M, Choe H. 2020. SARS-CoV-2 spike-protein D614G mutation increases virion spike density and infectivity. Nat Commun 11:6013. doi:10.1038/s41467-020-19808-4. - DOI - PMC - PubMed
-
- Laporte M, Raeymaekers V, Van Berwaer R, Vandeput J, Marchand-Casas I, Thibaut HJ, Van Looveren D, Martens K, Hoffmann M, Maes P, Pöhlmann S, Naesens L, Stevaert A. 2021. The SARS-CoV-2 and other human coronavirus spike proteins are fine-tuned towards temperature and proteases of the human airways. PLoS Pathog 17:e1009500. doi:10.1371/journal.ppat.1009500. - DOI - PMC - PubMed
-
- Hou YJ, Chiba S, Halfmann P, Ehre C, Kuroda M, Dinnon KH, 3rd, Leist SR, Schafer A, Nakajima N, Takahashi K, Lee RE, Mascenik TM, Graham R, Edwards CE, Tse LV, Okuda K, Markmann AJ, Bartelt L, de Silva A, Margolis DM, Boucher RC, Randell SH, Suzuki T, Gralinski LE, Kawaoka Y, Baric RS. 2020. SARS-CoV-2 D614G variant exhibits efficient replication ex vivo and transmission in vivo. Science 370:1464–1468. doi:10.1126/science.abe8499. - DOI - PMC - PubMed
-
- Zhou B, Thao TTN, Hoffmann D, Taddeo A, Ebert N, Labroussaa F, Pohlmann A, King J, Steiner S, Kelly JN, Portmann J, Halwe NJ, Ulrich L, Trueb BS, Fan X, Hoffmann B, Wang L, Thomann L, Lin X, Stalder H, Pozzi B, de Brot S, Jiang N, Cui D, Hossain J, Wilson MM, Keller MW, Stark TJ, Barnes JR, Dijkman R, Jores J, Benarafa C, Wentworth DE, Thiel V, Beer M. 2021. SARS-CoV-2 spike D614G change enhances replication and transmission. Nature 592:122–127. doi:10.1038/s41586-021-03361-1. - DOI - PubMed
-
- Saito A, Irie T, Suzuki R, Maemura T, Nasser H, Uriu K, Kosugi Y, Shirakawa K, Sadamasu K, Kimura I, Ito J, Wu J, Iwatsuki-Horimoto K, Ito M, Yamayoshi S, Loeber S, Tsuda M, Wang L, Ozono S, Butlertanaka EP, Tanaka YL, Shimizu R, Shimizu K, Yoshimatsu K, Kawabata R, Sakaguchi T, Tokunaga K, Yoshida I, Asakura H, Nagashima M, Kazuma Y, Nomura R, Horisawa Y, Yoshimura K, Takaori-Kondo A, Imai M, Tanaka S, Nakagawa S, Ikeda T, Fukuhara T, Kawaoka Y, Sato K, Genotype to Phenotype Japan Consortium . 2022. Enhanced fusogenicity and pathogenicity of SARS-CoV-2 Delta P681R mutation. Nature 602:300–306. doi:10.1038/s41586-021-04266-9. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
