

Mechanism of client selection by the protein quality-control factor UBE2O
- PMID: 35915257
- PMCID: PMC9526450
- DOI: 10.1038/s41594-022-00807-6
Mechanism of client selection by the protein quality-control factor UBE2O
Abstract
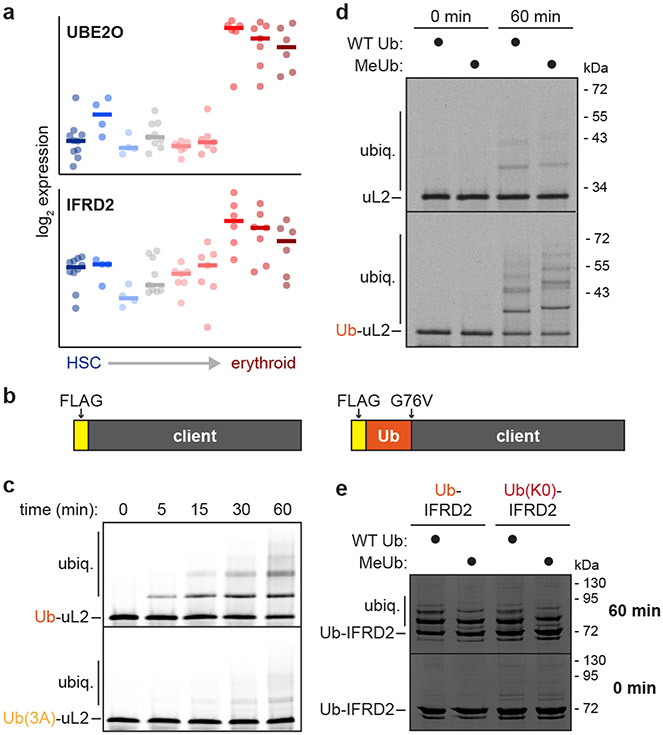
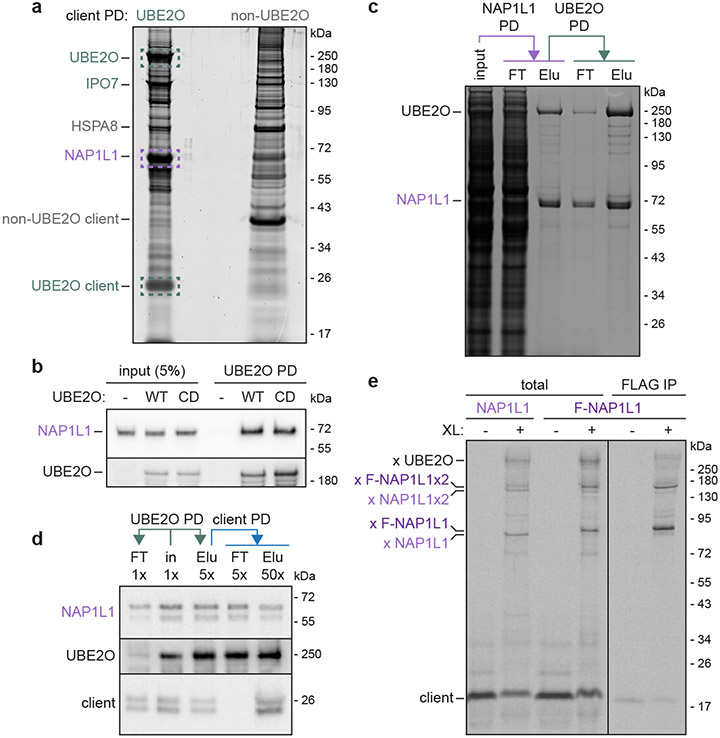
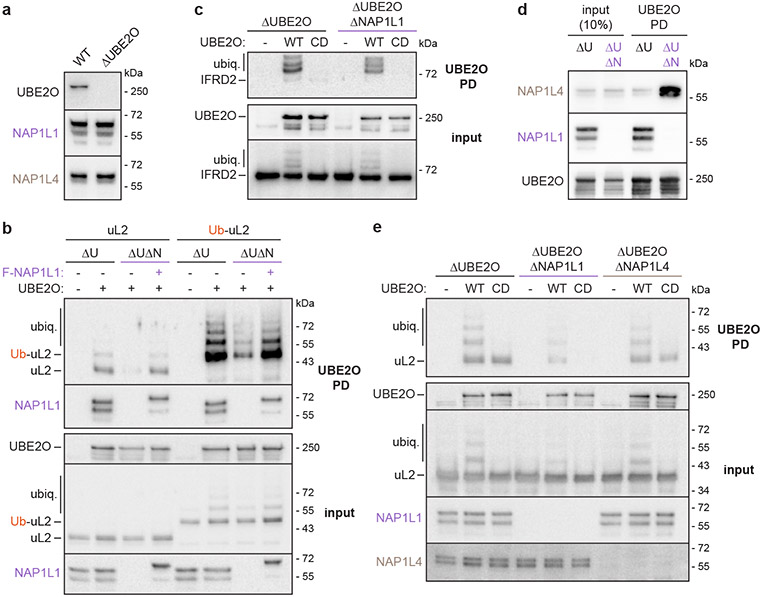
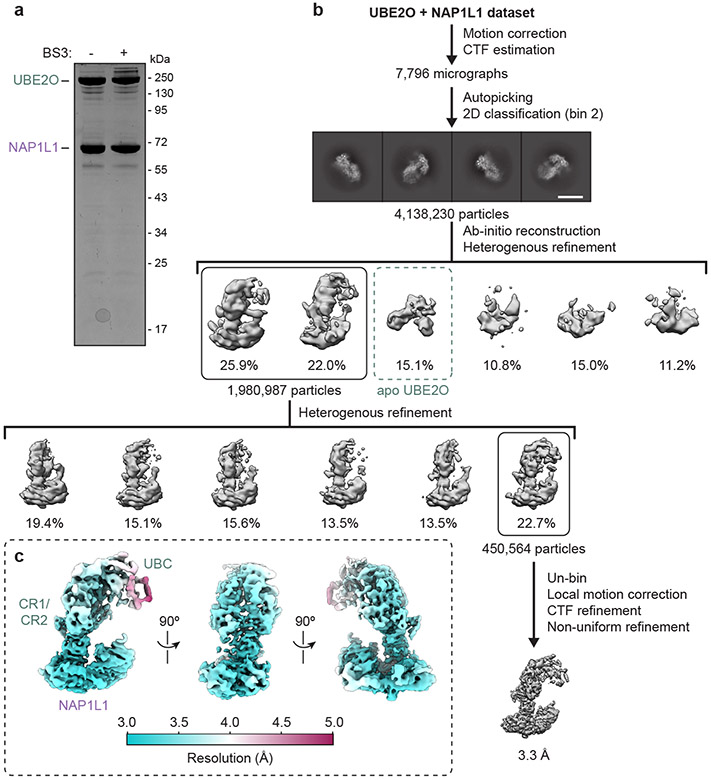
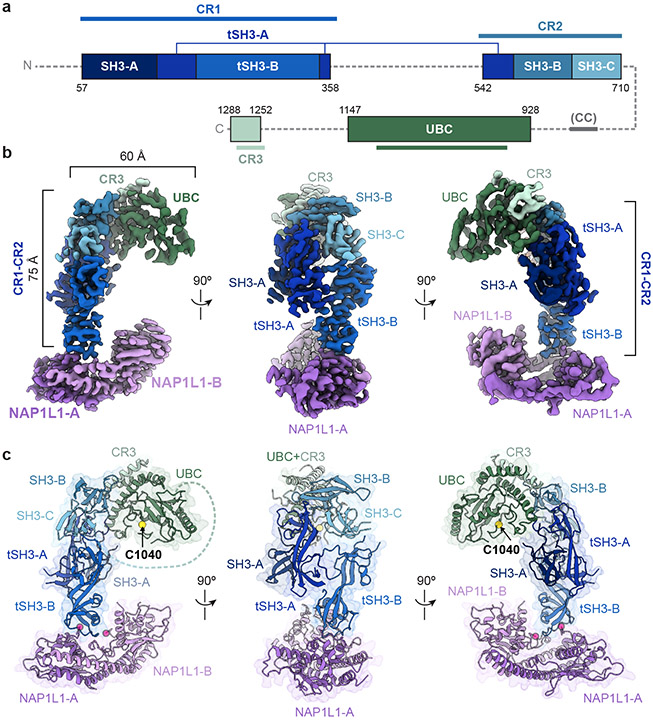
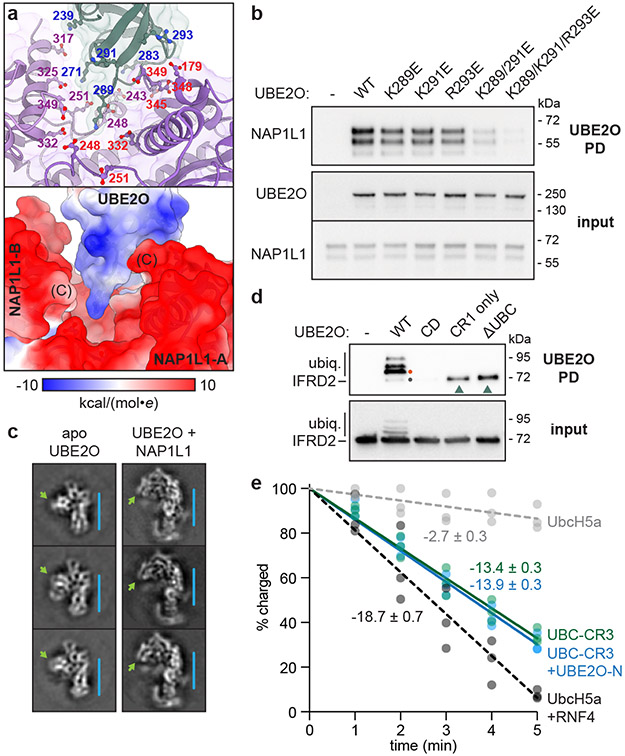
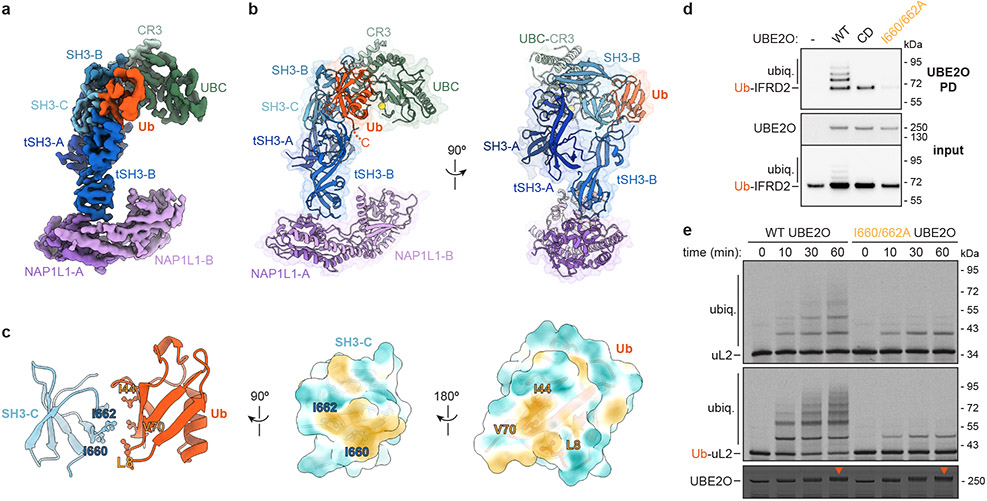
The E2/E3 enzyme UBE2O ubiquitylates diverse clients to mediate important processes, including targeting unassembled 'orphan' proteins for quality control and clearing ribosomes during erythropoiesis. How quality-control factors, such as UBE2O, select clients on the basis of heterogeneous features is largely unknown. Here, we show that UBE2O client selection is regulated by ubiquitin binding and a cofactor, NAP1L1. Attaching a single ubiquitin onto a client enhances UBE2O binding and multi-mono-ubiquitylation. UBE2O also repurposes the histone chaperone NAP1L1 as an adapter to recruit a subset of clients. Cryo-EM structures of human UBE2O in complex with NAP1L1 reveal a malleable client recruitment interface that is autoinhibited by the intrinsically reactive UBC domain. Adding a ubiquitylated client identifies a distinct ubiquitin-binding SH3-like domain required for client selection. Our findings reveal how multivalency and a feed-forward mechanism drive the selection of protein quality-control clients.
© 2022. The Author(s), under exclusive licence to Springer Nature America, Inc.
Figures
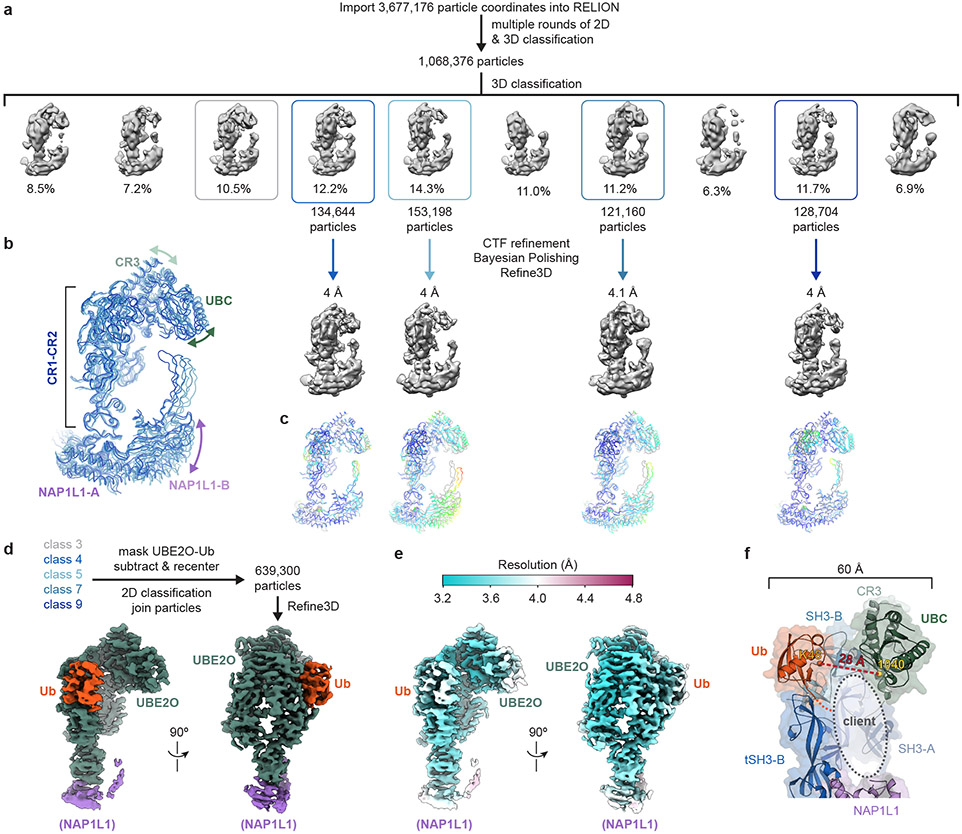
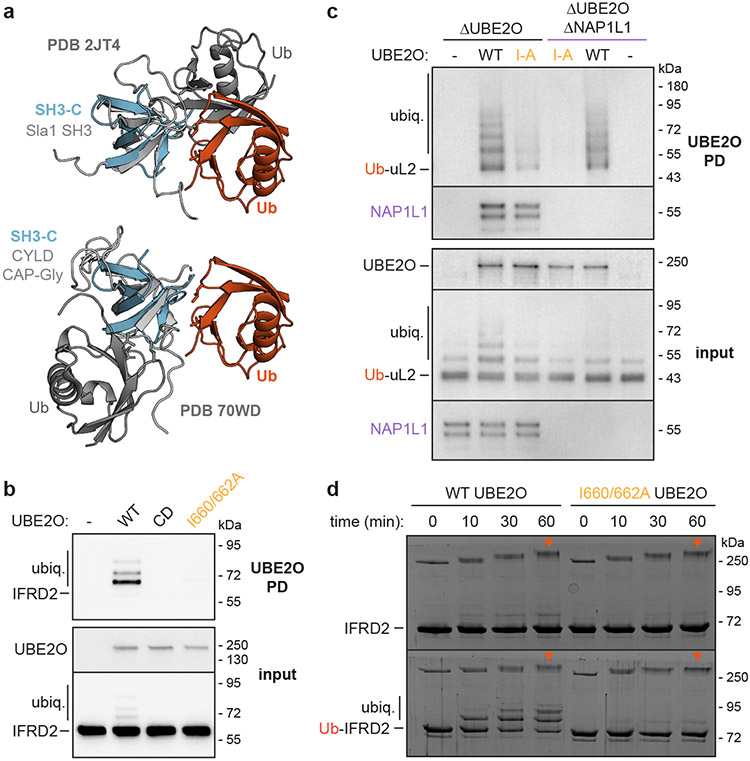
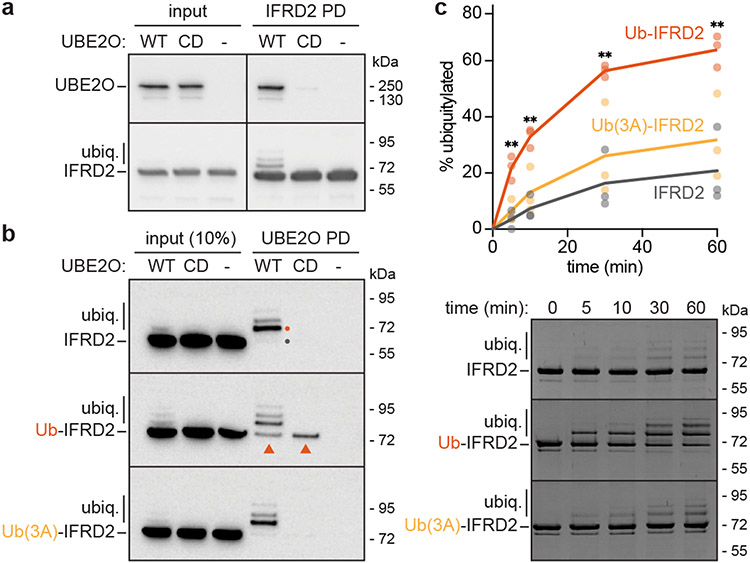
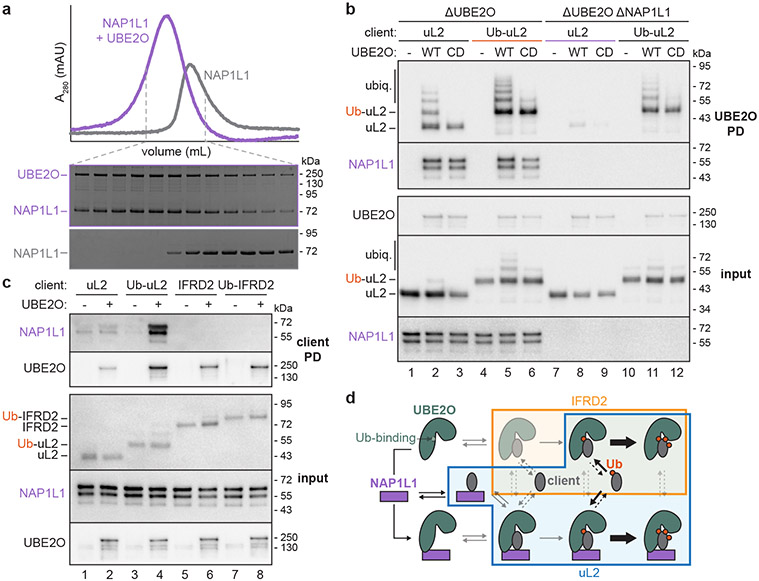
Similar articles
-
Structural insights into the biochemical mechanism of the E2/E3 hybrid enzyme UBE2O.Structure. 2025 Feb 6;33(2):274-288.e4. doi: 10.1016/j.str.2024.12.002. Epub 2024 Dec 30. Structure. 2025. PMID: 39740670
-
Ubiquitin-conjugating enzyme UBE2O regulates cellular clock function by promoting the degradation of the transcription factor BMAL1.J Biol Chem. 2018 Jul 20;293(29):11296-11309. doi: 10.1074/jbc.RA117.001432. Epub 2018 Jun 5. J Biol Chem. 2018. PMID: 29871923 Free PMC article.
-
Diverse roles of the E2/E3 hybrid enzyme UBE2O in the regulation of protein ubiquitination, cellular functions, and disease onset.FEBS J. 2019 Jun;286(11):2018-2034. doi: 10.1111/febs.14708. Epub 2018 Dec 4. FEBS J. 2019. PMID: 30468556 Review.
-
UBE2O is a quality control factor for orphans of multiprotein complexes.Science. 2017 Aug 4;357(6350):472-475. doi: 10.1126/science.aan0178. Science. 2017. PMID: 28774922 Free PMC article.
-
E2-E3 ubiquitin enzyme pairing - partnership in provoking or mitigating cancers.Biochim Biophys Acta Rev Cancer. 2022 Mar;1877(2):188679. doi: 10.1016/j.bbcan.2022.188679. Epub 2022 Jan 22. Biochim Biophys Acta Rev Cancer. 2022. PMID: 35074437 Review.
Cited by
-
Noncanonical assembly, neddylation and chimeric cullin-RING/RBR ubiquitylation by the 1.8 MDa CUL9 E3 ligase complex.Nat Struct Mol Biol. 2024 Jul;31(7):1083-1094. doi: 10.1038/s41594-024-01257-y. Epub 2024 Apr 11. Nat Struct Mol Biol. 2024. PMID: 38605244 Free PMC article.
-
CircDCAF8 promotes the progression of hepatocellular carcinoma through miR-217/NAP1L1 Axis, and induces angiogenesis and regorafenib resistance via exosome-mediated transfer.J Transl Med. 2024 May 30;22(1):517. doi: 10.1186/s12967-024-05233-4. J Transl Med. 2024. PMID: 38816735 Free PMC article.
-
A New Role of NAP1L1 in Megakaryocytes and Human Platelets.Int J Mol Sci. 2022 Nov 24;23(23):14694. doi: 10.3390/ijms232314694. Int J Mol Sci. 2022. PMID: 36499021 Free PMC article.
-
The Emerging Role and Mechanism of E2/E3 Hybrid Enzyme UBE2O in Human Diseases.Biomedicines. 2025 Apr 29;13(5):1082. doi: 10.3390/biomedicines13051082. Biomedicines. 2025. PMID: 40426910 Free PMC article. Review.
References
-
- Berleth ES & Pickart CM Mechanism of Ubiquitin Conjugating Enzyme E2-230K: Catalysis Involving a Thiol Relay? Biochemistry 35, 1664–1671 (1996). - PubMed
Methods References
-
- Shimizu Y & Ueda T PURE technology. Methods Mol Biol 607, 11–21 (2010). - PubMed
-
- Feng Q & Shao S In vitro reconstitution of translational arrest pathways. Methods 137, 20–36 (2018). - PubMed
-
- Punjani A, Rubinstein JL, Fleet DJ & Brubaker MA cryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat Methods 14, 290–296 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
