

Differential role of neuronal glucose and PFKFB3 in memory formation during development
- PMID: 35916383
- PMCID: PMC9474594
- DOI: 10.1002/glia.24248
Differential role of neuronal glucose and PFKFB3 in memory formation during development
Abstract
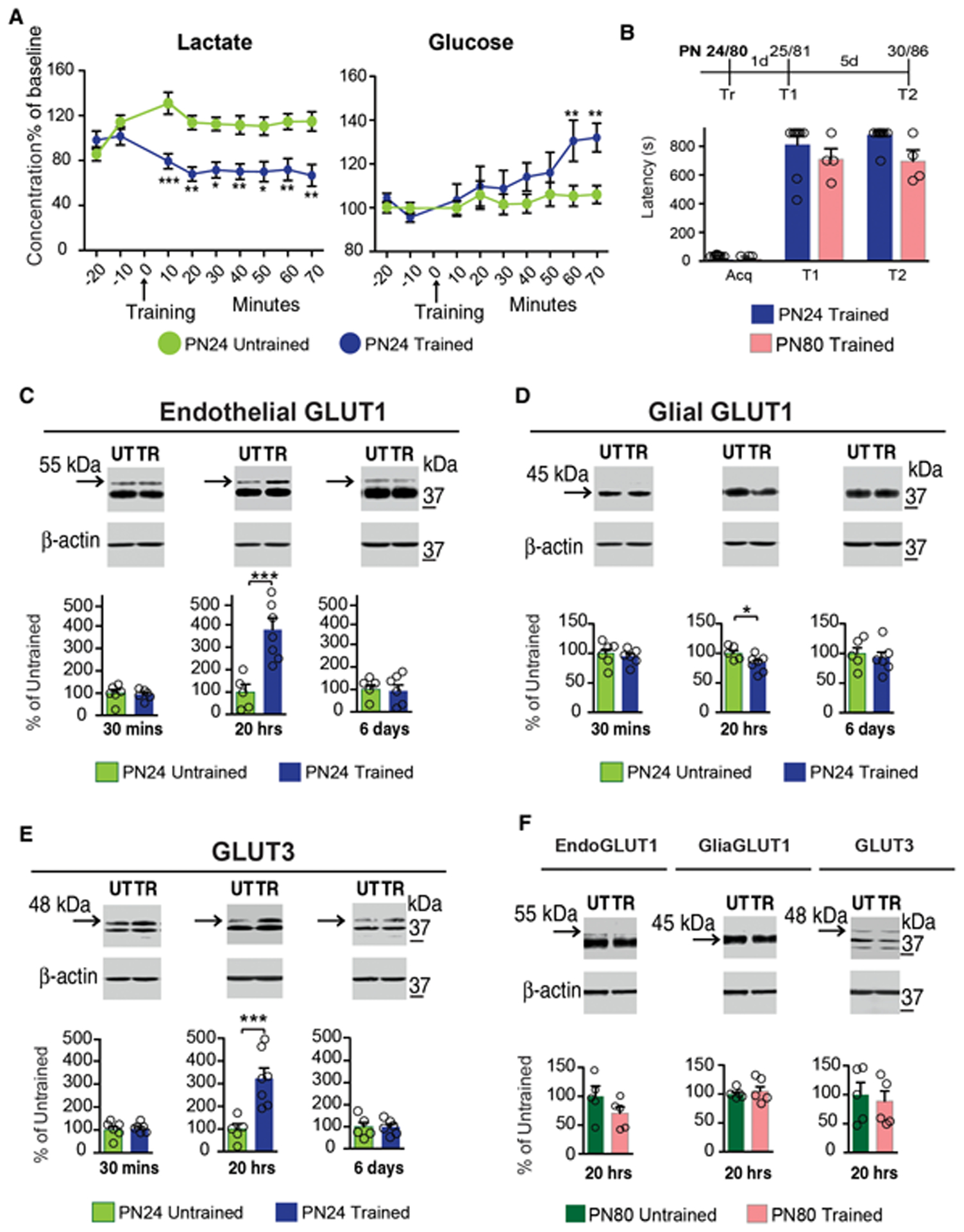
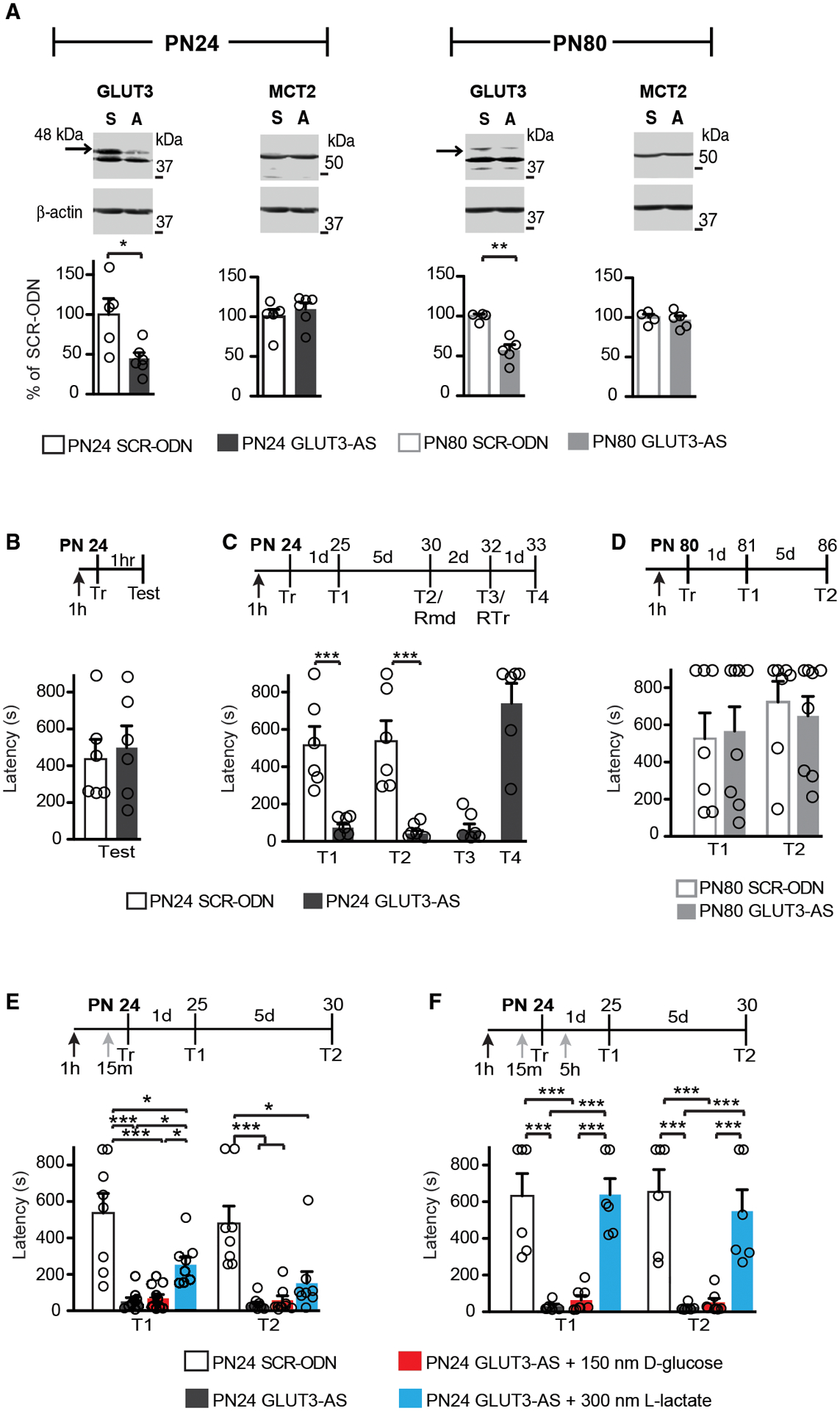
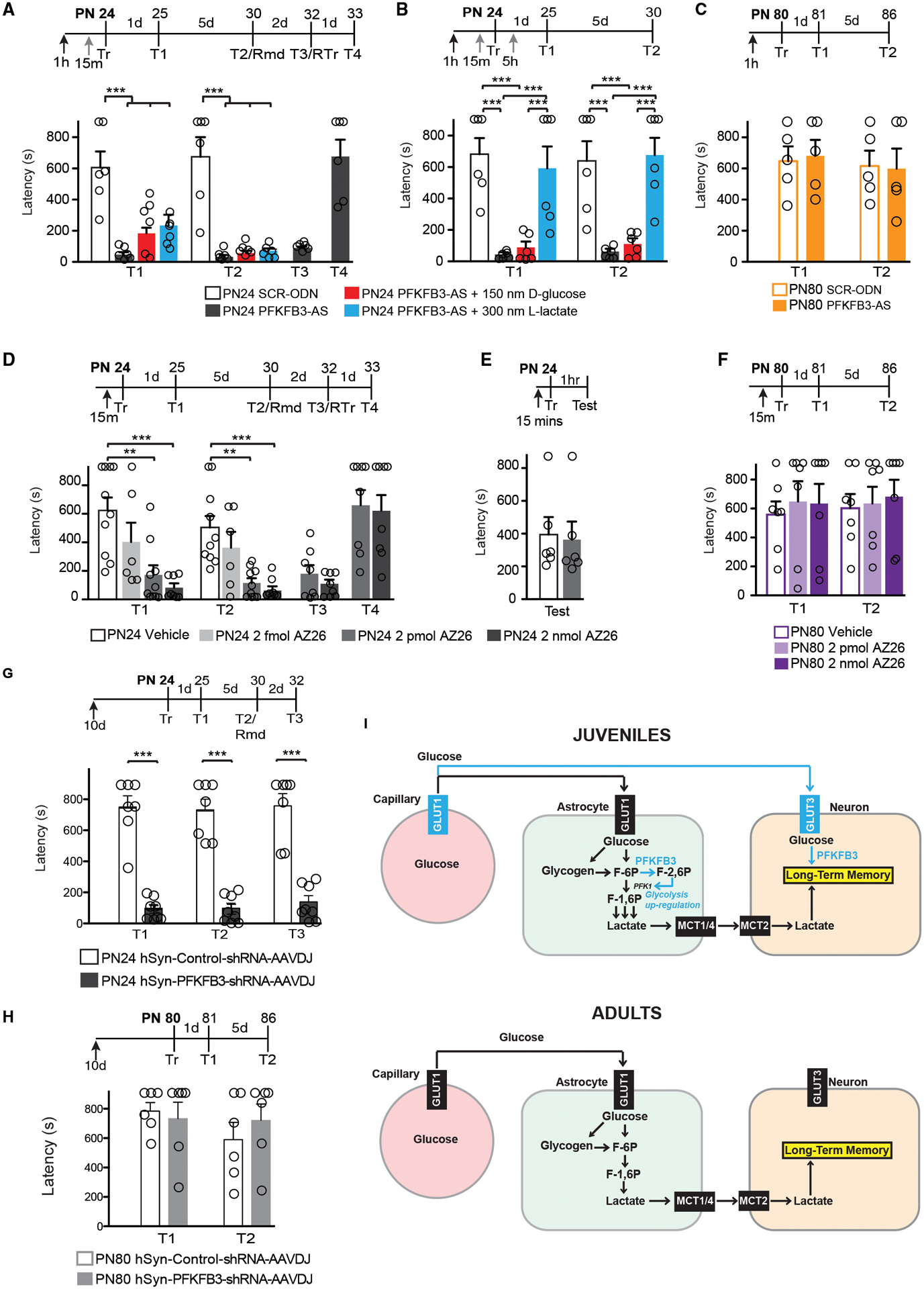
The consumption of glucose in the brain peaks during late childhood; yet, whether and how glucose metabolism is differentially regulated in the brain during childhood compared to adulthood remains to be understood. In particular, it remains to be determined how glucose metabolism is involved in behavioral activations such as learning. Here we show that, compared to adult, the juvenile rat hippocampus has significantly higher mRNA levels of several glucose metabolism enzymes belonging to all glucose metabolism pathways, as well as higher levels of the monocarboxylate transporters MCT1 and MCT4 and the glucose transporters endothelial-GLUT1 and GLUT3 proteins. Furthermore, relative to adults, long-term episodic memory formation in juvenile animals requires significantly higher rates of aerobic glycolysis and astrocytic-neuronal lactate coupling in the hippocampus. Only juvenile but not adult long-term memory formation recruits GLUT3, neuronal 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 3 (PFKFB3) and more efficiently engages glucose in the hippocampus. Hence, compared to adult, the juvenile hippocampus distinctively regulates glucose metabolism pathways, and formation of long-term memory in juveniles involves differential neuronal glucose metabolism mechanisms.
Keywords: GLUT3; PFKFB3; development; glucose; hippocampus; lactate; memory.
© 2022 Wiley Periodicals LLC.
Conflict of interest statement
DECLARATION OF INTEREST
The authors declare no competing interests.
Figures




Similar articles
-
PFKFB3-mediated glycolysis is involved in reactive astrocyte proliferation after oxygen-glucose deprivation/reperfusion and is regulated by Cdh1.Neurochem Int. 2015 Dec;91:26-33. doi: 10.1016/j.neuint.2015.10.006. Epub 2015 Oct 21. Neurochem Int. 2015. PMID: 26498254
-
6-Phosphofructo-2-kinase/fructose-2,6-biphosphatase 3 and 4: A pair of valves for fine-tuning of glucose metabolism in human cancer.Mol Metab. 2019 Feb;20:1-13. doi: 10.1016/j.molmet.2018.11.013. Epub 2018 Dec 5. Mol Metab. 2019. PMID: 30553771 Free PMC article. Review.
-
Monocarboxylate transporter 1 promotes classical microglial activation and pro-inflammatory effect via 6-phosphofructo-2-kinase/fructose-2, 6-biphosphatase 3.J Neuroinflammation. 2019 Nov 28;16(1):240. doi: 10.1186/s12974-019-1648-4. J Neuroinflammation. 2019. PMID: 31779643 Free PMC article.
-
Hypoxic preconditioning up-regulates glucose transport activity and glucose transporter (GLUT1 and GLUT3) gene expression after acute anoxic exposure in the cultured rat hippocampal neurons and astrocytes.Brain Res. 2008 May 23;1211:22-9. doi: 10.1016/j.brainres.2005.04.029. Brain Res. 2008. PMID: 18474279
-
Treatment against glucose-dependent cancers through metabolic PFKFB3 targeting of glycolytic flux.Cancer Metastasis Rev. 2022 Jun;41(2):447-458. doi: 10.1007/s10555-022-10027-5. Epub 2022 Apr 14. Cancer Metastasis Rev. 2022. PMID: 35419769 Review.
Cited by
-
A temporal cortex cell atlas highlights gene expression dynamics during human brain maturation.Nat Genet. 2024 Dec;56(12):2718-2730. doi: 10.1038/s41588-024-01990-6. Epub 2024 Nov 20. Nat Genet. 2024. PMID: 39567748 Free PMC article.
-
Cross-talk between energy and redox metabolism in astrocyte-neuron functional cooperation.Essays Biochem. 2023 Mar 3;67(1):17-26. doi: 10.1042/EBC20220075. Essays Biochem. 2023. PMID: 36805653 Free PMC article. Review.
-
Aerobic glycolysis is the predominant means of glucose metabolism in neuronal somata, which protects against oxidative damage.Nat Neurosci. 2023 Dec;26(12):2081-2089. doi: 10.1038/s41593-023-01476-4. Epub 2023 Nov 23. Nat Neurosci. 2023. PMID: 37996529
-
Weak neuronal glycolysis sustains cognition and organismal fitness.Nat Metab. 2024 Jul;6(7):1253-1267. doi: 10.1038/s42255-024-01049-0. Epub 2024 May 24. Nat Metab. 2024. PMID: 38789798 Free PMC article.
-
Brain energy metabolism: A roadmap for future research.J Neurochem. 2024 May;168(5):910-954. doi: 10.1111/jnc.16032. Epub 2024 Jan 6. J Neurochem. 2024. PMID: 38183680 Free PMC article. Review.
References
-
- Allaman I, and Magistretti PJ (2013). Brain Energy Metabolism. In Fundamental Neuroscience: Fourth Edition, p.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
