

Inactivated genotype 1a, 2a and 3a HCV vaccine candidates induced broadly neutralising antibodies in mice
- PMID: 35918103
- PMCID: PMC9933178
- DOI: 10.1136/gutjnl-2021-326323
Inactivated genotype 1a, 2a and 3a HCV vaccine candidates induced broadly neutralising antibodies in mice
Abstract
Objective: A prophylactic vaccine is needed to control the HCV epidemic, with genotypes 1-3 causing >80% of worldwide infections. Vaccine development is hampered by HCV heterogeneity, viral escape including protection of conserved neutralising epitopes and suboptimal efficacy of HCV cell culture systems. We developed cell culture-based inactivated genotype 1-3 HCV vaccine candidates to present natively folded envelope proteins to elicit neutralising antibodies.
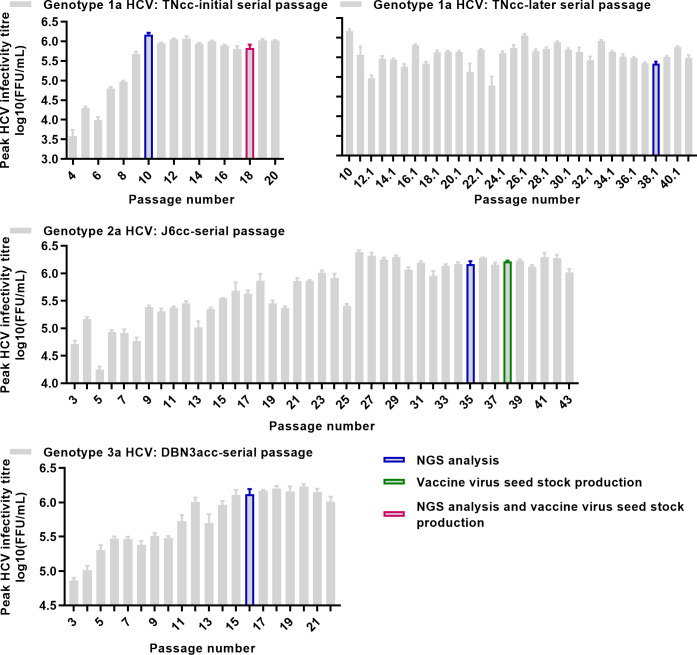
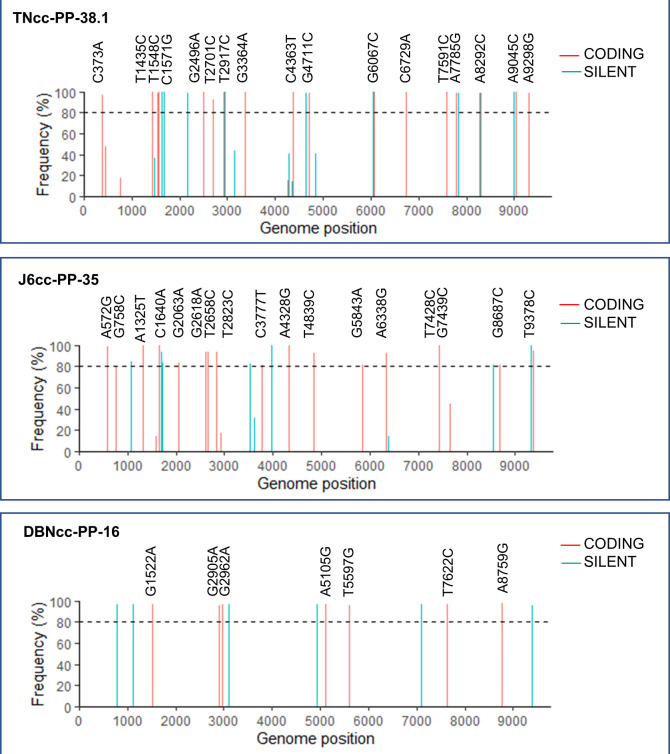
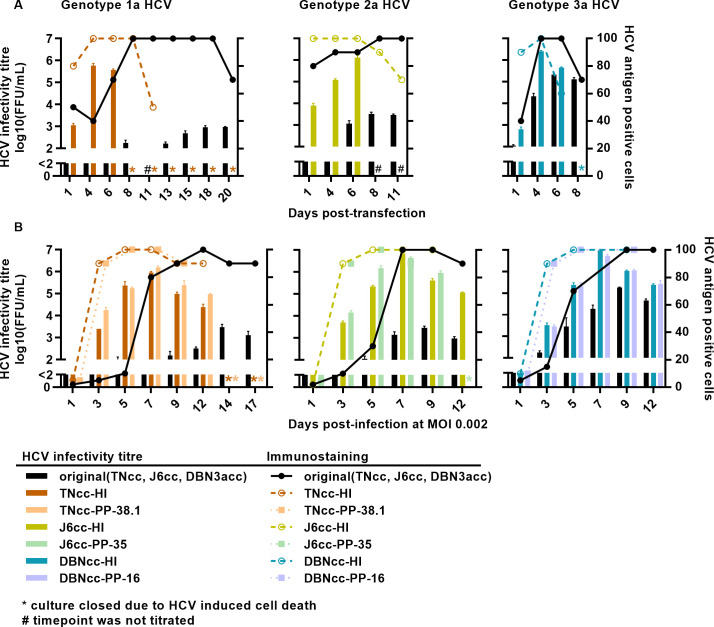
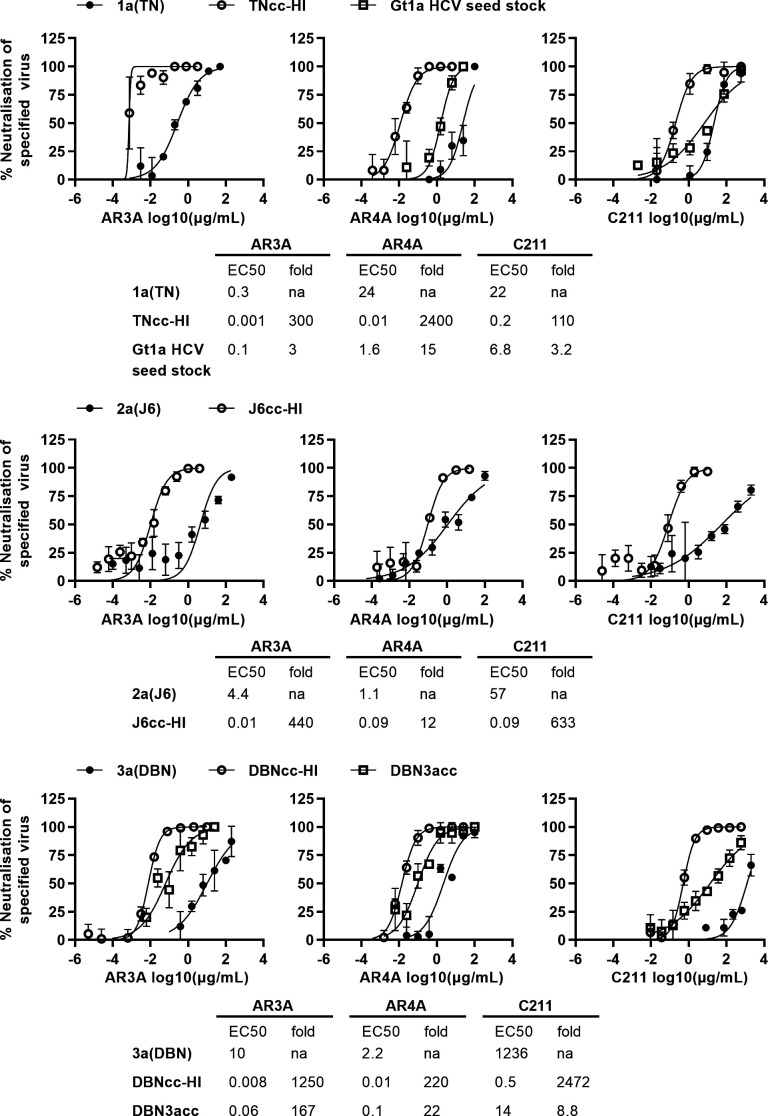
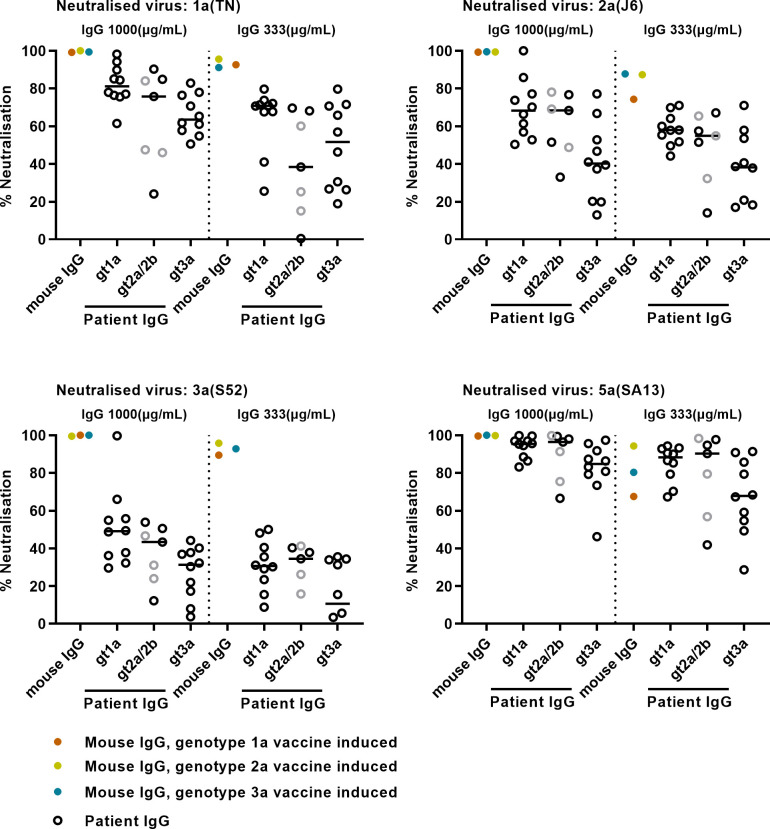
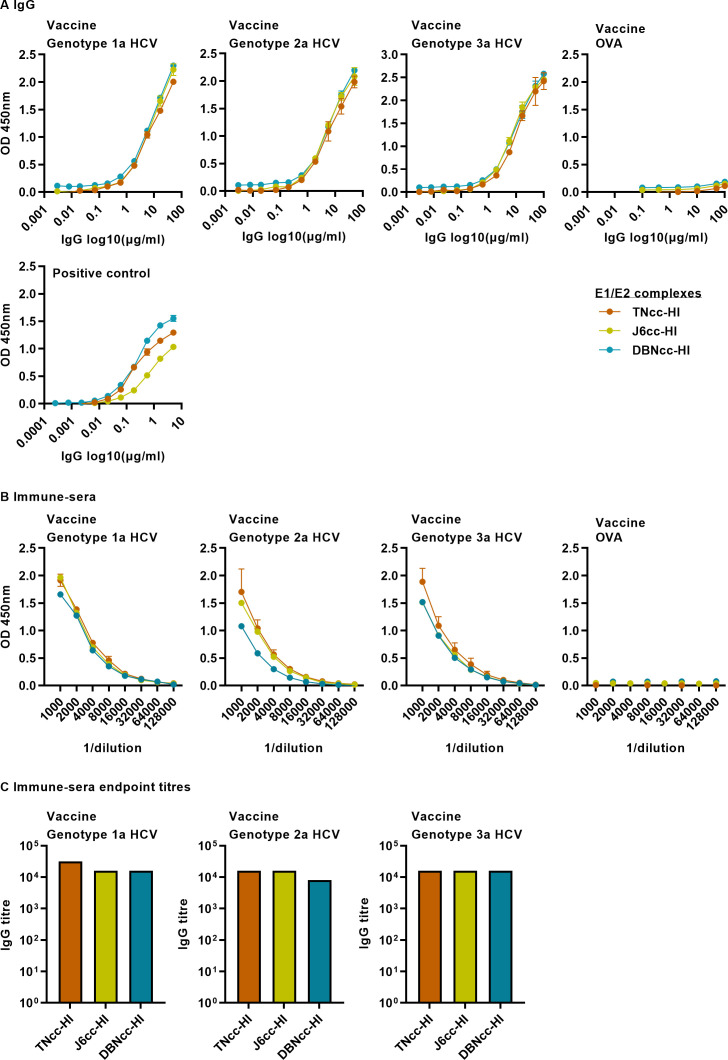
Design: High-yield genotype 1a, 2a and 3a HCV were developed by serial passage of TNcc, J6cc and DBN3acc in Huh7.5 cells and engineering of acquired mutations detected by next-generation sequencing. Neutralising epitope exposure was determined in cell-based neutralisation assays using human monoclonal antibodies AR3A and AR4A, and polyclonal antibody C211. BALB/c mice were immunised with processed and inactivated genotype 1a, 2a or 3a viruses using AddaVax, a homologue of the licenced adjuvant MF-59. Purified mouse and patient serum IgG were assayed for neutralisation capacity; mouse IgG and immune-sera were assayed for E1/E2 binding.
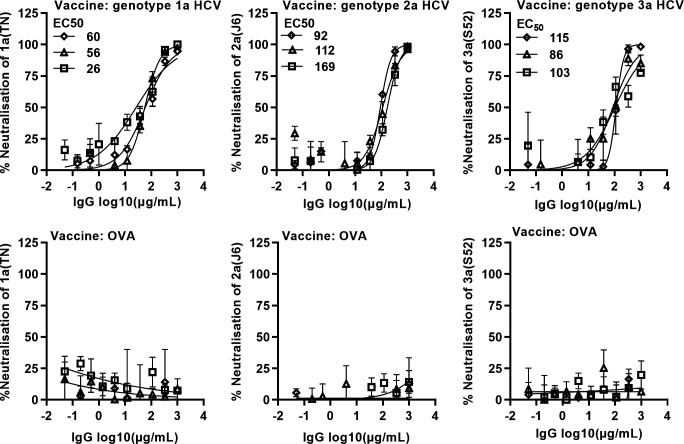
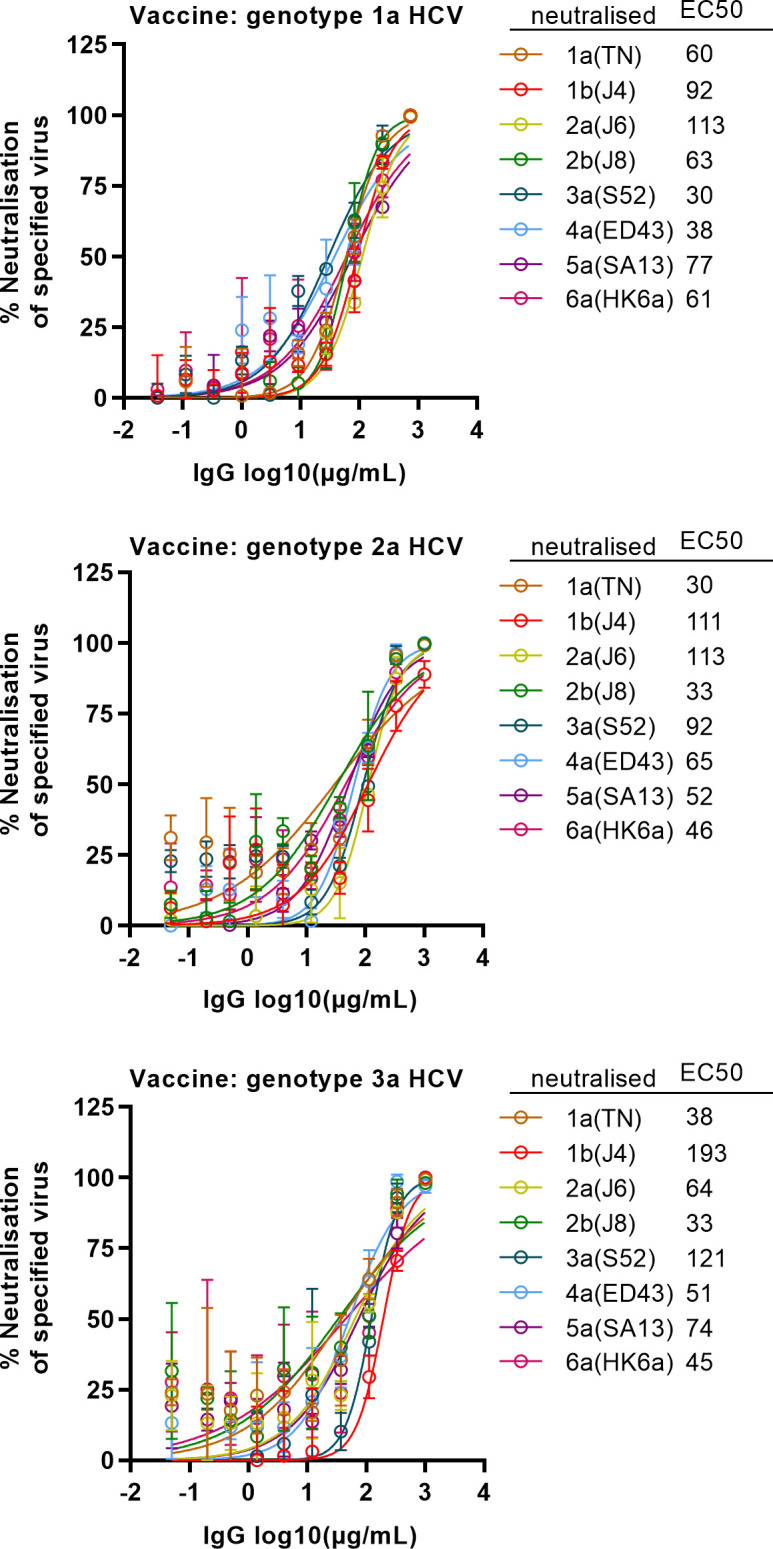
Results: Compared with the original viruses, high-yield viruses had up to ~1000 fold increased infectivity titres (peak titres: 6-7 log10 focus-forming units (FFU)/mL) and up to ~2470 fold increased exposure of conserved neutralising epitopes. Vaccine-induced IgG broadly neutralised genotype 1-6 HCV (EC50: 30-193 µg/mL; mean 71 µg/mL), compared favourably with IgG from chronically infected patients, and bound genotype 1-3 E1/E2; immune-sera endpoint titres reached up to 32 000.
Conclusion: High-yield genotype 1-3 HCV could be developed as basis for inactivated vaccine candidates inducing broadly neutralising antibodies in mice supporting further preclinical development.
Keywords: HCV; chronic viral hepatitis; hepatitis C; immune response; molecular biology.
© Author(s) (or their employer(s)) 2023. Re-use permitted under CC BY-NC. No commercial re-use. See rights and permissions. Published by BMJ.
Conflict of interest statement
Competing interests: None declared.
Figures
References
-
- Hepatitis C. Available: https://www.who.int/news-room/fact-sheets/detail/hepatitis-c [Accessed April 29, 2022].
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous