

Genomic architecture of adaptive radiation and hybridization in Alpine whitefish
- PMID: 35918341
- PMCID: PMC9345977
- DOI: 10.1038/s41467-022-32181-8
Genomic architecture of adaptive radiation and hybridization in Alpine whitefish
Abstract
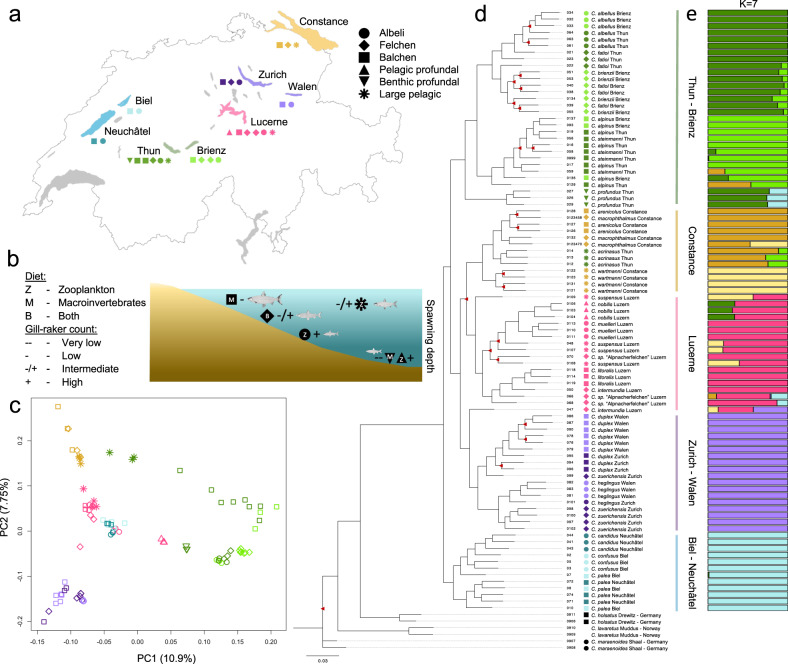
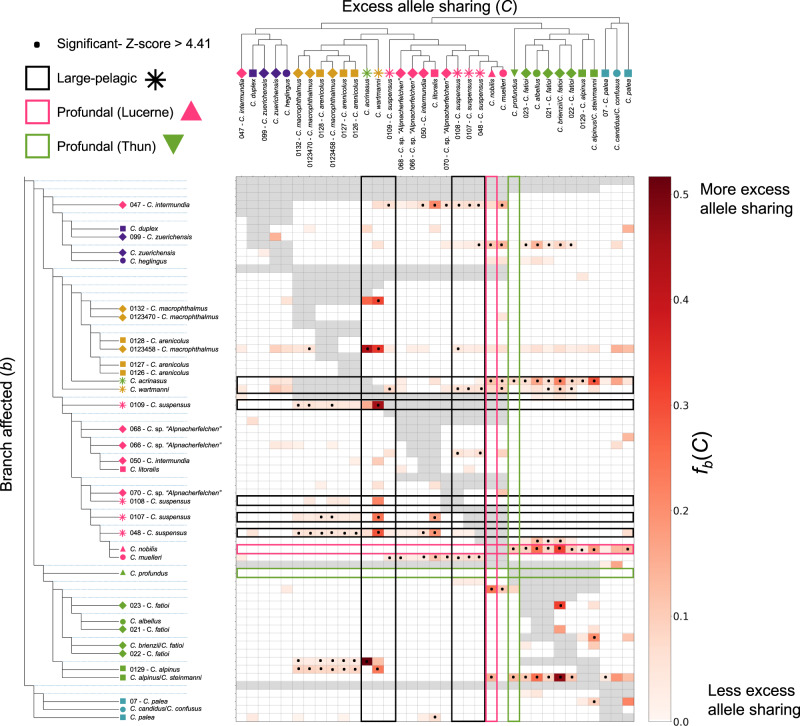
Adaptive radiations represent some of the most remarkable explosions of diversification across the tree of life. However, the constraints to rapid diversification and how they are sometimes overcome, particularly the relative roles of genetic architecture and hybridization, remain unclear. Here, we address these questions in the Alpine whitefish radiation, using a whole-genome dataset that includes multiple individuals of each of the 22 species belonging to six ecologically distinct ecomorph classes across several lake-systems. We reveal that repeated ecological and morphological diversification along a common environmental axis is associated with both genome-wide allele frequency shifts and a specific, larger effect, locus, associated with the gene edar. Additionally, we highlight the possible role of introgression between species from different lake-systems in facilitating the evolution and persistence of species with unique trait combinations and ecology. These results highlight the importance of both genome architecture and secondary contact with hybridization in fuelling adaptive radiation.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Nosil, P. Ecological Speciation (Oxford University Press, 2012).
-
- Schluter, D. The Ecology of Adaptive Radiation (OUP Oxford, 2000).
-
- Gavrilets, S. Fitness Landscapes and the Origin of Species (Princeton University Press, 2004).
