

BTApep-TAT peptide inhibits ADP-ribosylation of BORIS to induce DNA damage in cancer
- PMID: 35918747
- PMCID: PMC9344678
- DOI: 10.1186/s12943-022-01621-w
BTApep-TAT peptide inhibits ADP-ribosylation of BORIS to induce DNA damage in cancer
Abstract
Background: Brother of regulator of imprinted sites (BORIS) is expressed in most cancers and often associated with short survival and poor prognosis in patients. BORIS inhibits apoptosis and promotes proliferation of cancer cells. However, its mechanism of action has not been elucidated, and there is no known inhibitor of BORIS.
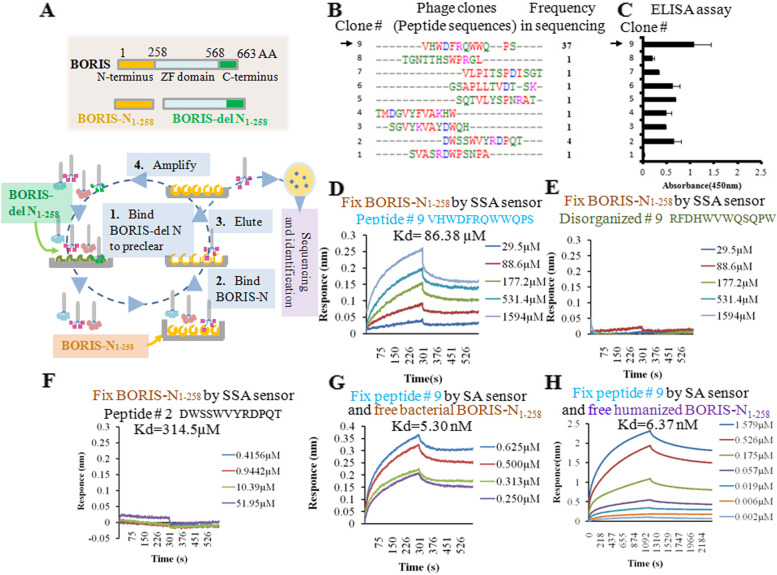
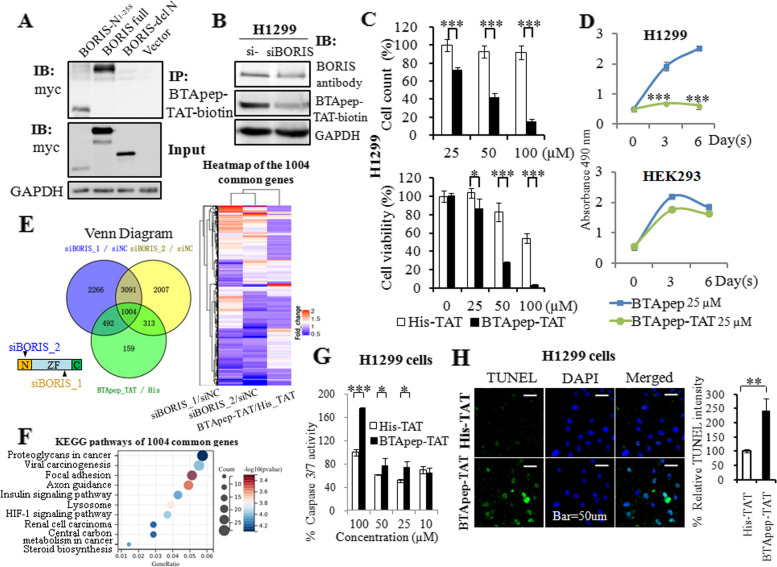
Methods: A phage display library was used to find the BORIS inhibitory peptides and BTApep-TAT was identified. The RNA sequencing profile of BTApep-TAT-treated H1299 cells was compared with that of BORIS-knockdown cells. Antitumor activity of BTApep-TAT was evaluated in a non-small cell lung cancer (NSCLC) xenograft mouse model. BTApep-TAT was also used to investigate the post-translational modification (PTM) of BORIS and the role of BORIS in DNA damage repair. Site-directed mutants of BORIS were constructed and used for investigating PTM and the function of BORIS.
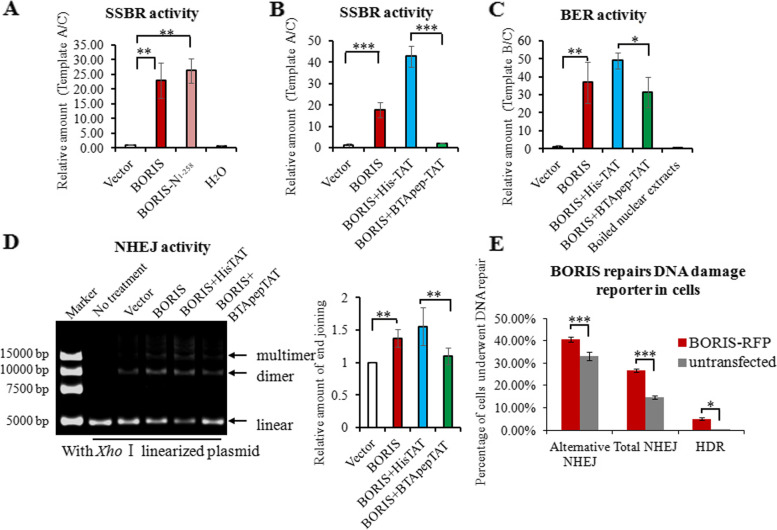
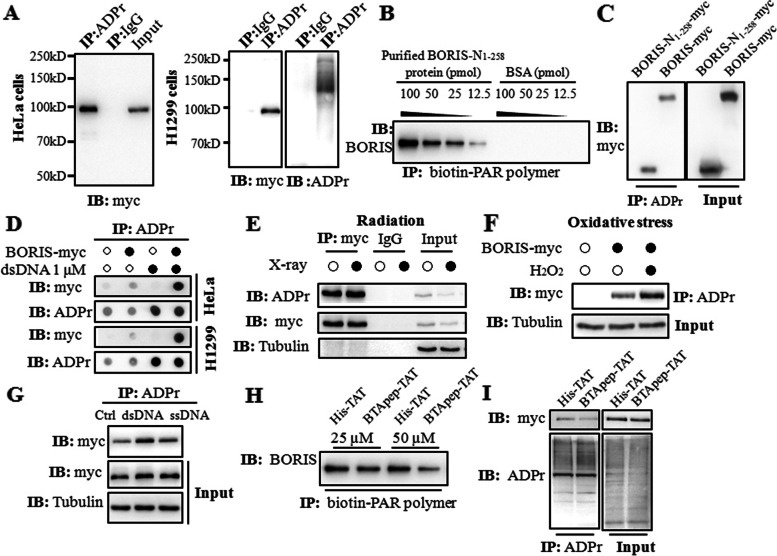
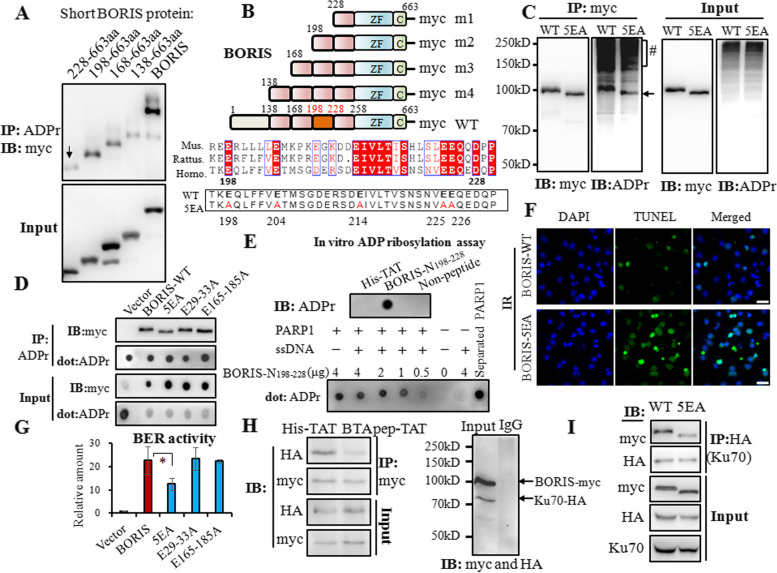
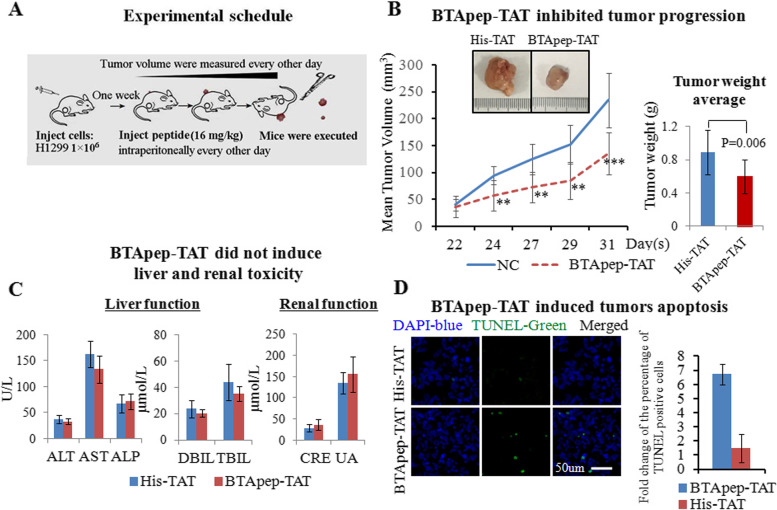
Results: BTApep-TAT induced DNA damage in cancer cells and suppressed NSCLC xenograft tumor progression. Investigation of the mechanism of action of BTApep-TAT demonstrated that BORIS underwent ADP ribosylation upon double- or single-strand DNA damage. Substitution of five conserved glutamic acid (E) residues with alanine residues (A) between amino acids (AAs) 198 and 228 of BORIS reduced its ADP ribosylation. Inhibition of ADP ribosylation of BORIS by a site-specific mutation or by BTApep-TAT treatment blocked its interaction with Ku70 and impaired the function of BORIS in DNA damage repair.
Conclusions: The present study identified an inhibitor of BORIS, highlighted the importance of ADP ribosylation of BORIS, and revealed a novel function of BORIS in DNA damage repair. The present work provides a practical method for the future screening or optimization of drugs targeting BORIS.
Keywords: ADP-ribosylation; BORIS; DNA damage; Non-small cell lung cancer; Targeted peptide.
© 2022. The Author(s).
Conflict of interest statement
We declare that we have no financial and personal relationships with other people or organizations that can inappropriately influence our work, and there is no professional or other personal interest of any nature or kind in any product, service and/or company that could be construed as influencing the position presented in, or the review of, the manuscript entitled.
Figures
Similar articles
-
Brother of regulator of imprinted sites inhibits cisplatin-induced DNA damage in non-small cell lung cancer.Oncol Lett. 2020 Nov;20(5):251. doi: 10.3892/ol.2020.12114. Epub 2020 Sep 17. Oncol Lett. 2020. PMID: 32994814 Free PMC article.
-
Brother of the regulator of the imprinted site (BORIS) variant subfamily 6 is a novel target of lung cancer stem-like cell immunotherapy.PLoS One. 2017 Mar 1;12(3):e0171460. doi: 10.1371/journal.pone.0171460. eCollection 2017. PLoS One. 2017. PMID: 28248963 Free PMC article.
-
The function and regulation of ADP-ribosylation in the DNA damage response.Biochem Soc Trans. 2023 Jun 28;51(3):995-1008. doi: 10.1042/BST20220749. Biochem Soc Trans. 2023. PMID: 37171085 Free PMC article.
-
Serine ADP-ribosylation in DNA-damage response regulation.Curr Opin Genet Dev. 2021 Dec;71:106-113. doi: 10.1016/j.gde.2021.07.005. Epub 2021 Jul 31. Curr Opin Genet Dev. 2021. PMID: 34340015 Review.
-
ADP-ribosylhydrolases: from DNA damage repair to COVID-19.J Zhejiang Univ Sci B. 2021 Jan 15;22(1):21-30. doi: 10.1631/jzus.B2000319. J Zhejiang Univ Sci B. 2021. PMID: 33448184 Free PMC article. Review.
Cited by
-
Cancer testis antigens: Emerging therapeutic targets leveraging genomic instability in cancer.Mol Ther Oncol. 2024 Jan 26;32(1):200768. doi: 10.1016/j.omton.2024.200768. eCollection 2024 Mar 21. Mol Ther Oncol. 2024. PMID: 38596293 Free PMC article. Review.
-
CTCF and Its Multi-Partner Network for Chromatin Regulation.Cells. 2023 May 10;12(10):1357. doi: 10.3390/cells12101357. Cells. 2023. PMID: 37408191 Free PMC article. Review.
-
Deciphering the endogenous SUMO-1 landscape: a novel combinatorial peptide enrichment strategy for global profiling and disease association.Chem Sci. 2024 Dec 26;16(6):2634-2647. doi: 10.1039/d4sc07379g. eCollection 2025 Feb 5. Chem Sci. 2024. PMID: 39802689 Free PMC article.
-
DNAzyme hydrogels specifically inhibit the NLRP3 pathway to prevent radiation-induced skin injury in mice.J Nanobiotechnology. 2025 Mar 22;23(1):238. doi: 10.1186/s12951-025-03147-x. J Nanobiotechnology. 2025. PMID: 40119386 Free PMC article.
-
SUMOylation-induced membrane localization of TRPV1 suppresses proliferation and migration in gastric cancer cells.Cell Commun Signal. 2024 Sep 30;22(1):465. doi: 10.1186/s12964-024-01850-0. Cell Commun Signal. 2024. PMID: 39350261 Free PMC article.
References
-
- Asano T, Hirohashi Y, Torigoe T, Mariya T, Horibe R, Kuroda T, Tabuchi Y, Saijo H, Yasuda K, Mizuuchi M, et al. Brother of the regulator of the imprinted site (BORIS) variant subfamily 6 is involved in cervical cancer stemness and can be a target of immunotherapy. Oncotarget. 2016;7:11223–11237. doi: 10.18632/oncotarget.7165. - DOI - PMC - PubMed
-
- Loukinov DI, Pugacheva E, Vatolin S, Pack SD, Moon H, Chernukhin I, Mannan P, Larsson E, Kanduri C, Vostrov AA, et al. BORIS, a novel male germ-line-specific protein associated with epigenetic reprogramming events, shares the same 11-zinc-finger domain with CTCF, the insulator protein involved in reading imprinting marks in the soma. Proc Natl Acad Sci U S A. 2002;99:6806–6811. doi: 10.1073/pnas.092123699. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
