

The second polar body contributes to the fate asymmetry in the mouse embryo
- PMID: 35919785
- PMCID: PMC9337984
- DOI: 10.1093/nsr/nwac003
The second polar body contributes to the fate asymmetry in the mouse embryo
Abstract
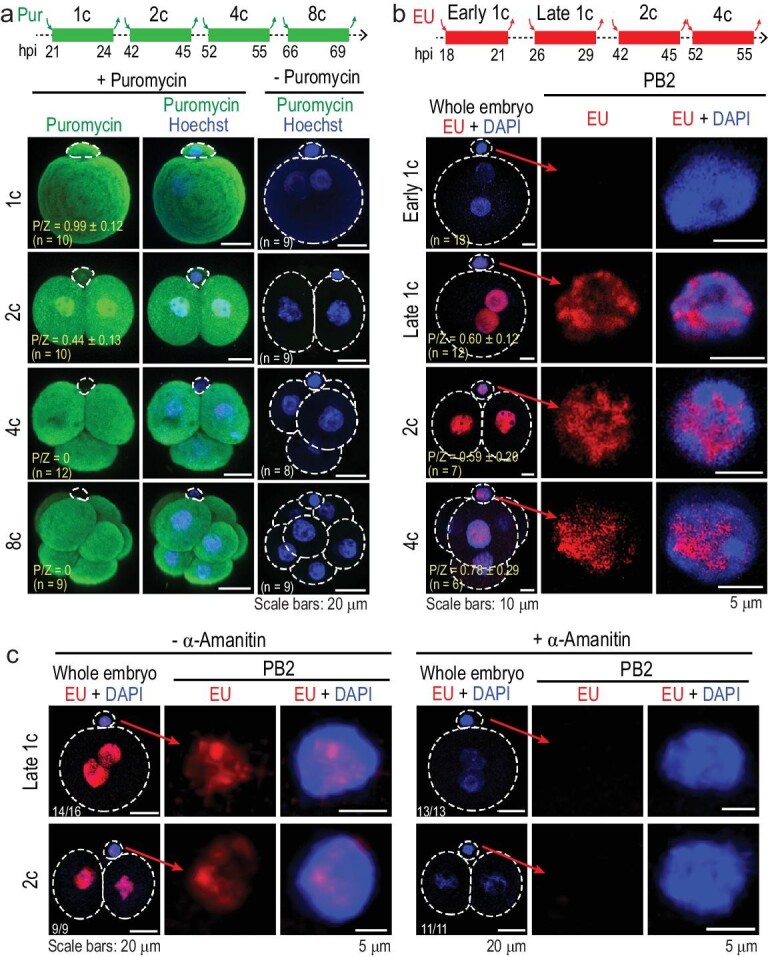
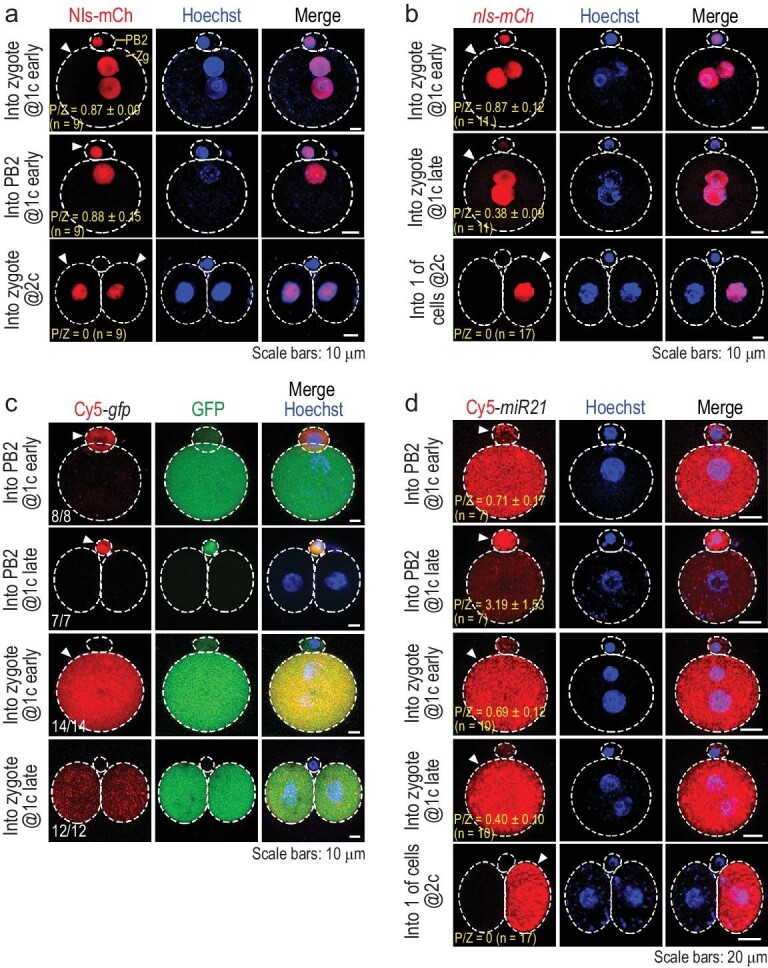
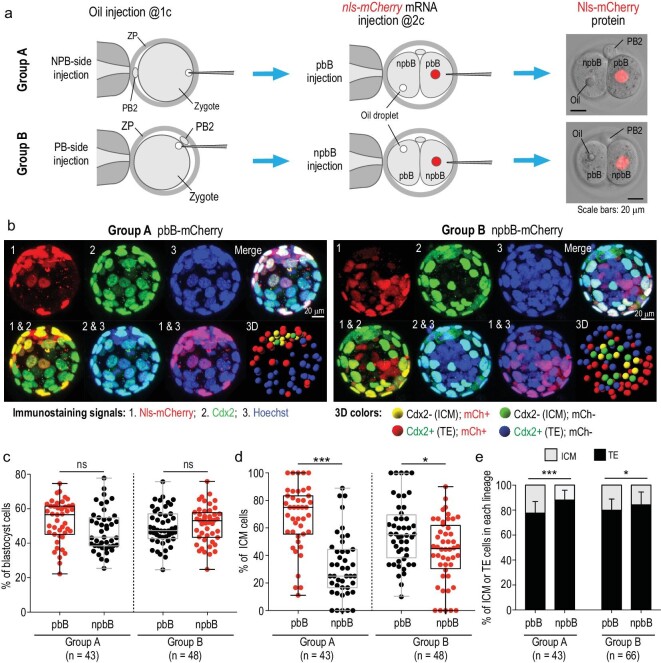
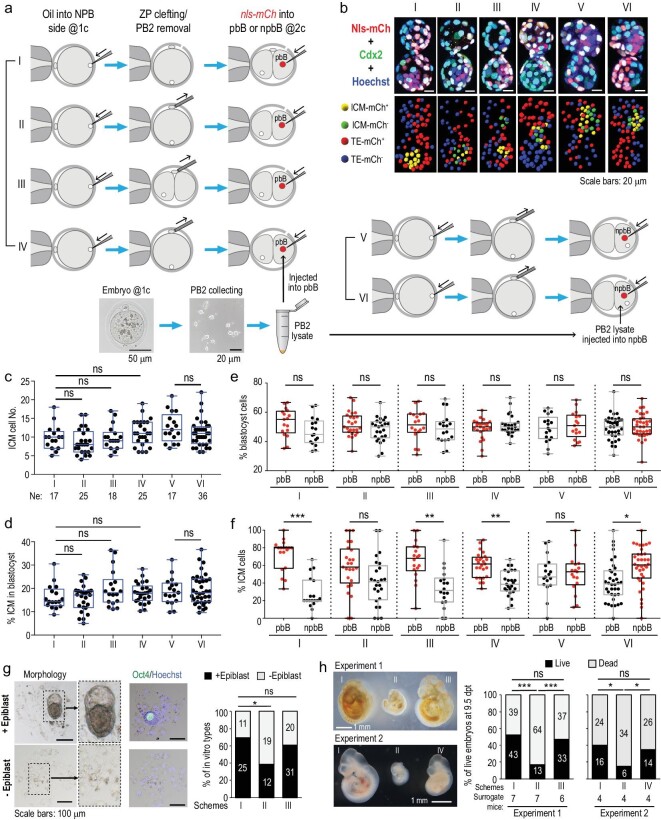
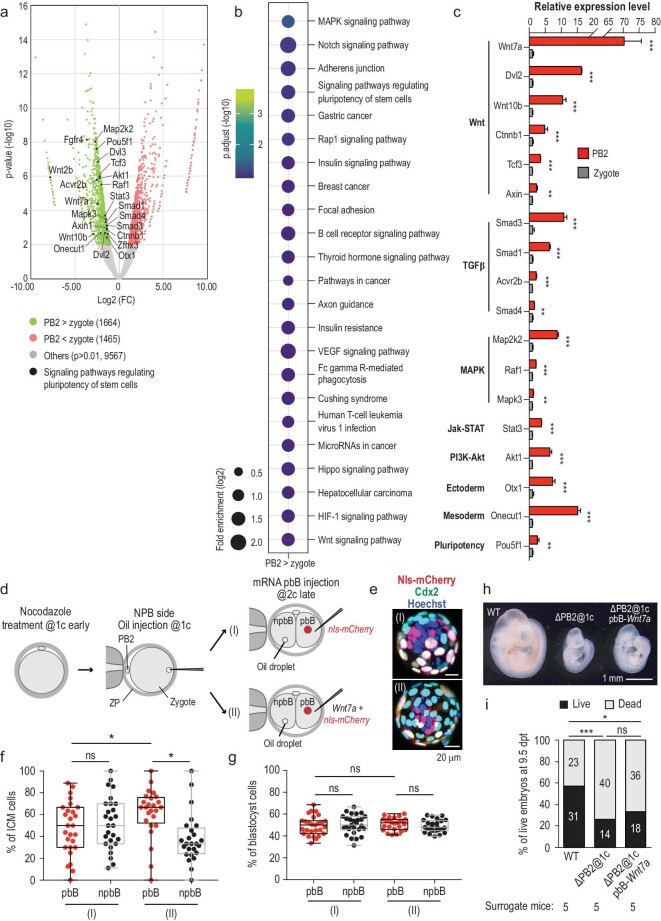
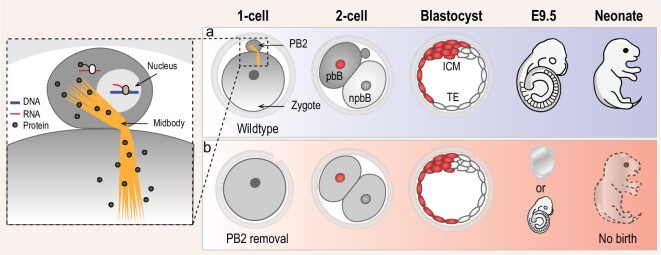
The polar bodies (PBs) are extruded microcells during oocyte meiosis and generally regarded as inessentials for embryonic development. Therefore, PBs have been widely used as important materials for pre-implantation genetic diagnosis in human. Here we report that the second PB (PB2) in the mouse zygote may play roles in cell-fate specification and post-implantation development. A subset of mRNAs encoding pluripotency-related factors are enriched in PB2. Nascent proteins may be synthesized in PB2 after fertilization and transport from PB2 to the zygote before the two-cell stage. The PB2-attached blastomere (pbB) at the two-cell stage, compared to the other blastomere (npbB), likely contributes more descendants to the inner cell mass (ICM) lineage in the blastocyst. Removal of PB2 from the zygote or transient blockage of material exchange between PB2 and the zygote by nocodazole treatment appears to cause a loss of the ICM fate bias of pbB. PB2 removal or nocodazole treatment also results in abnormal post-implantation development. Injection of PB2 lysate into pbB of PB2-removed two-cell-stage embryos may reset the cell-fate preference and rescue post-implantation development. Our data collectively suggest that PB2 would demarcate the earliest cell-fate asymmetry of the mouse zygote and be required for post-implantation development.
Keywords: asymmetry; cell fate; embryo; mouse; polar body.
© The Author(s) 2022. Published by Oxford University Press on behalf of China Science Publishing & Media Ltd.
Figures
Similar articles
-
Sequential comprehensive chromosome analysis on polar bodies, blastomeres and trophoblast: insights into female meiotic errors and chromosomal segregation in the preimplantation window of embryo development.Hum Reprod. 2013 Feb;28(2):509-18. doi: 10.1093/humrep/des394. Epub 2012 Nov 11. Hum Reprod. 2013. PMID: 23148203
-
Role of PB1 Midbody Remnant Creating Tethered Polar Bodies during Meiosis II.Genes (Basel). 2020 Nov 24;11(12):1394. doi: 10.3390/genes11121394. Genes (Basel). 2020. PMID: 33255457 Free PMC article.
-
Polar body analysis by array comparative genomic hybridization accurately predicts aneuploidies of maternal meiotic origin in cleavage stage embryos of women of advanced maternal age.Hum Reprod. 2013 May;28(5):1426-34. doi: 10.1093/humrep/det053. Epub 2013 Mar 10. Hum Reprod. 2013. PMID: 23477909
-
Epigenetic control of cell fate in mouse blastocysts: the role of covalent histone modifications and chromatin remodeling.Mol Reprod Dev. 2014 Feb;81(2):171-82. doi: 10.1002/mrd.22219. Epub 2013 Aug 13. Mol Reprod Dev. 2014. PMID: 23893501 Free PMC article. Review.
-
Morphogenetic tissue movement and the establishment of body plan during development from blastocyst to gastrula in the mouse.Bioessays. 2001 Jun;23(6):508-17. doi: 10.1002/bies.1070. Bioessays. 2001. PMID: 11385630 Review.
Cited by
-
Cellular mechanisms of monozygotic twinning: clues from assisted reproduction.Hum Reprod Update. 2024 Dec 1;30(6):692-705. doi: 10.1093/humupd/dmae022. Hum Reprod Update. 2024. PMID: 38996087 Free PMC article. Review.
-
Systematic evaluation of retroviral LTRs as cis-regulatory elements in mouse embryos.Cell Rep. 2024 Mar 26;43(3):113775. doi: 10.1016/j.celrep.2024.113775. Epub 2024 Feb 20. Cell Rep. 2024. PMID: 38381606 Free PMC article.
-
Efficient generation of all ESC-derived mice carrying a homozygous lethal mutation through eight-cell embryo injection.Sci China Life Sci. 2025 May;68(5):1524-1527. doi: 10.1007/s11427-024-2762-3. Epub 2025 Feb 13. Sci China Life Sci. 2025. PMID: 39960622 No abstract available.
-
Correlation of the position and status of the polar body from the fertilized oocyte to the euploid status of blastocysts.Front Genet. 2022 Sep 20;13:1006870. doi: 10.3389/fgene.2022.1006870. eCollection 2022. Front Genet. 2022. PMID: 36204310 Free PMC article.
-
Morphokinetic Behavior of the Second Polar Body in Human Zygotes as a Predictor for Embryonic Developmental Potential: An Exploratory Study Based on Time-Lapse Observation.Int J Mol Sci. 2025 Mar 29;26(7):3190. doi: 10.3390/ijms26073190. Int J Mol Sci. 2025. PMID: 40243997 Free PMC article.
References
-
- Nusslein-Volhard C. Determination of the embryonic axes of Drosophila. Dev Suppl 1991; 1: 1–10. - PubMed
LinkOut - more resources
Full Text Sources