

Tension-driven multi-scale self-organisation in human iPSC-derived muscle fibers
- PMID: 35920628
- PMCID: PMC9377800
- DOI: 10.7554/eLife.76649
Tension-driven multi-scale self-organisation in human iPSC-derived muscle fibers
Abstract
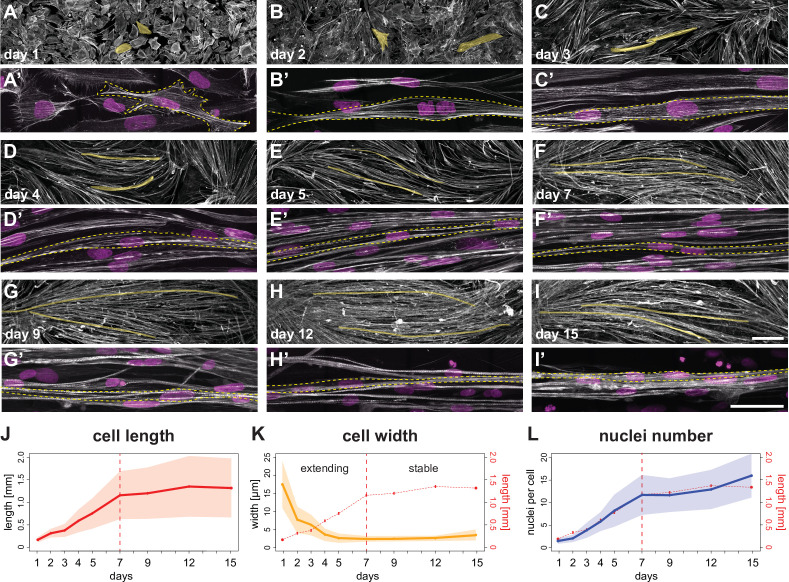
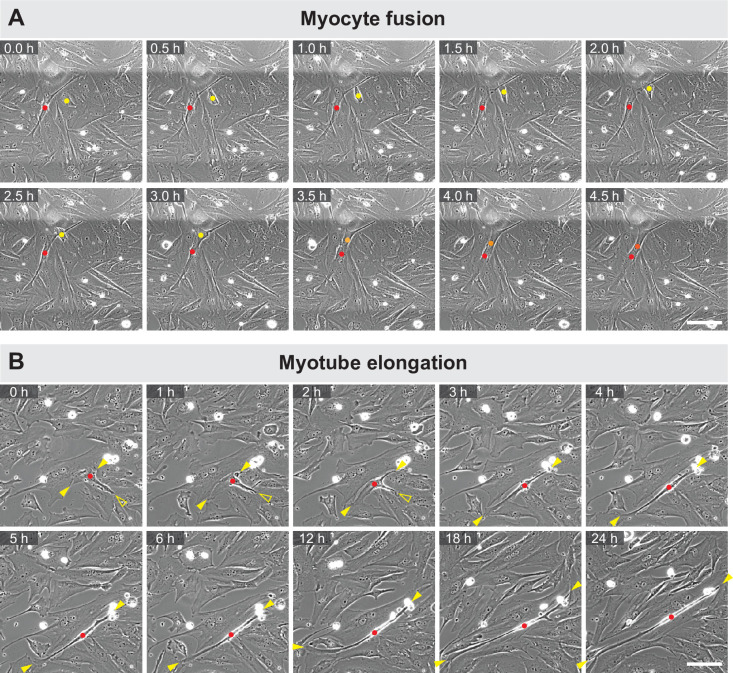
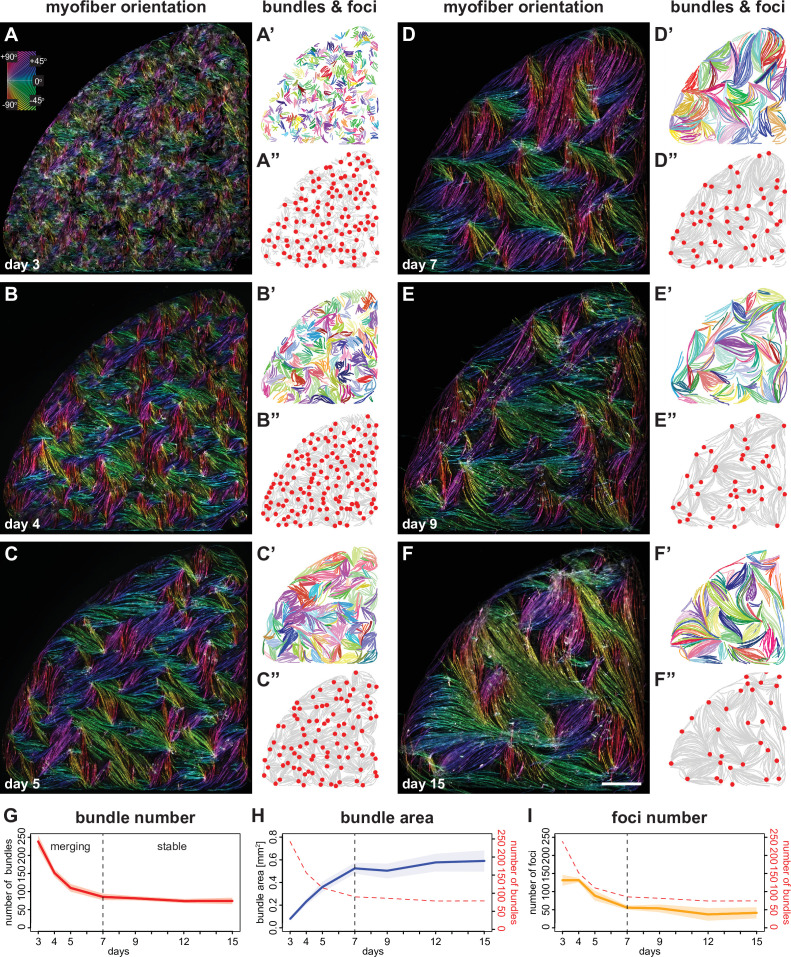
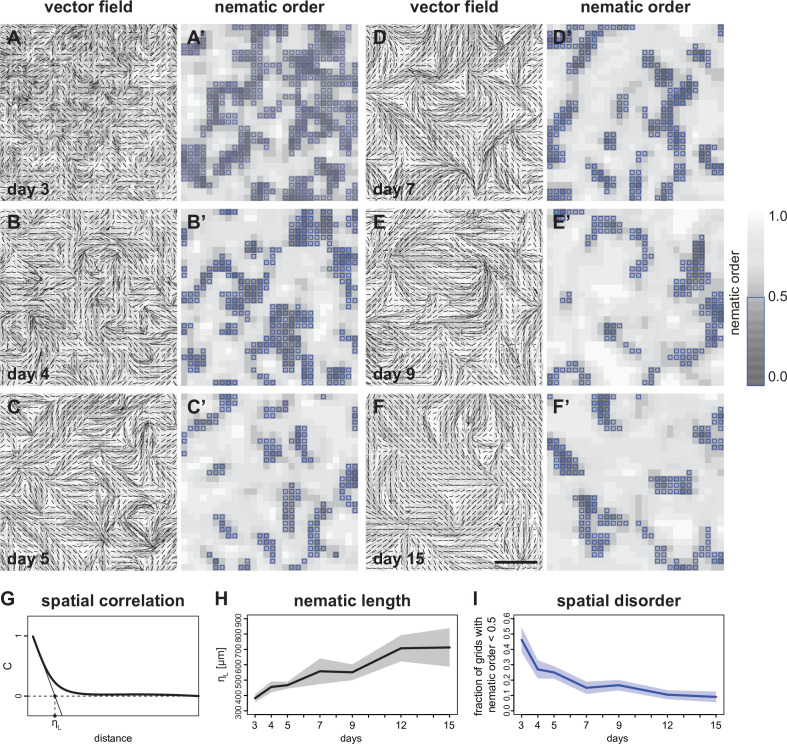
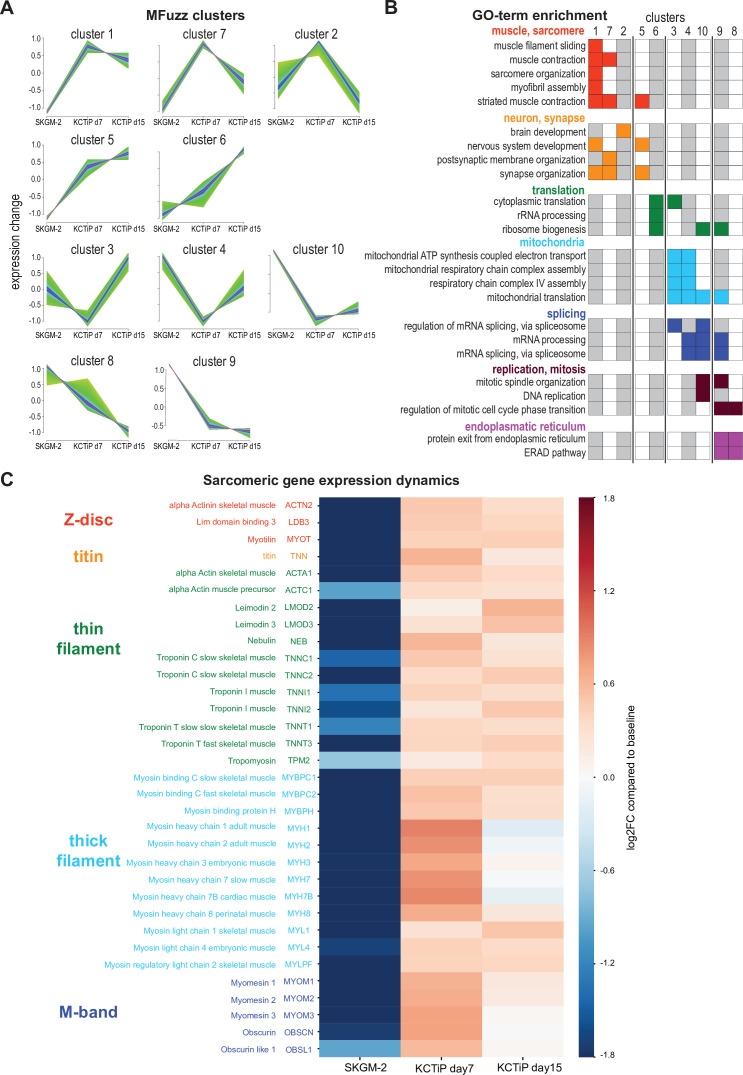
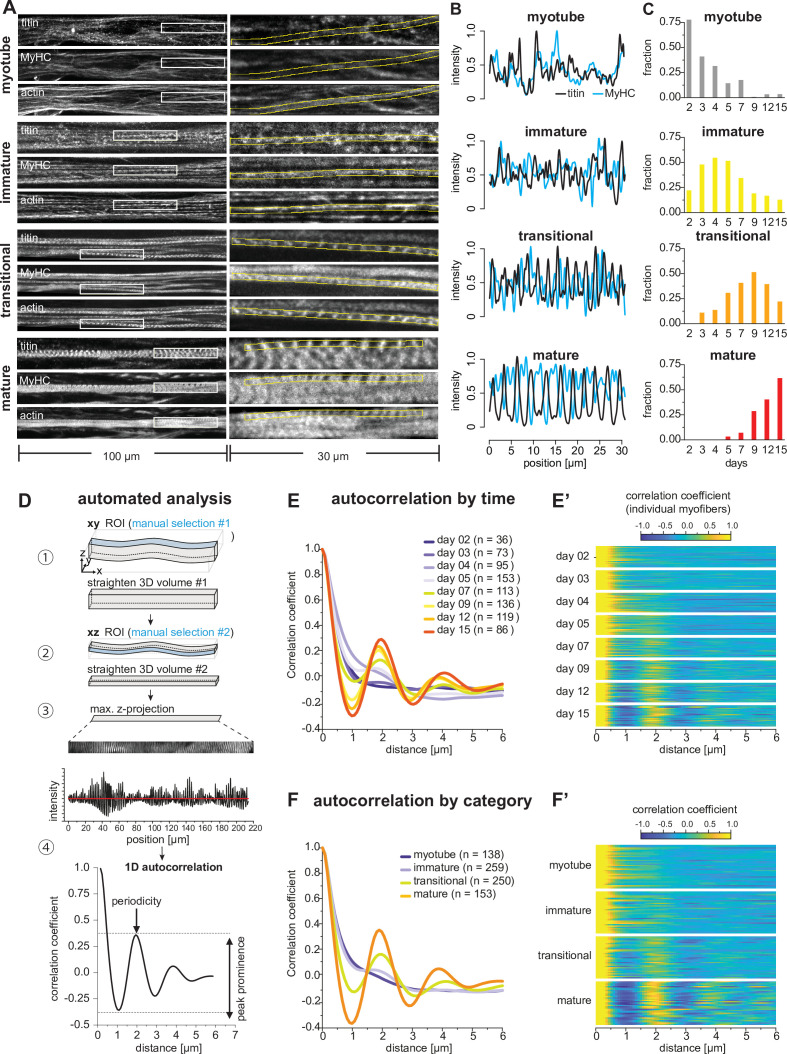
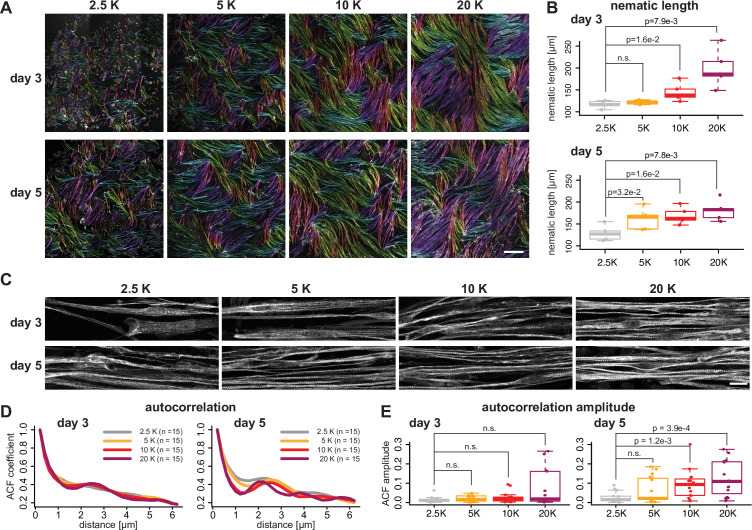
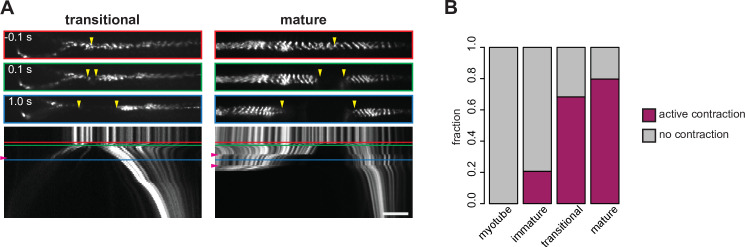
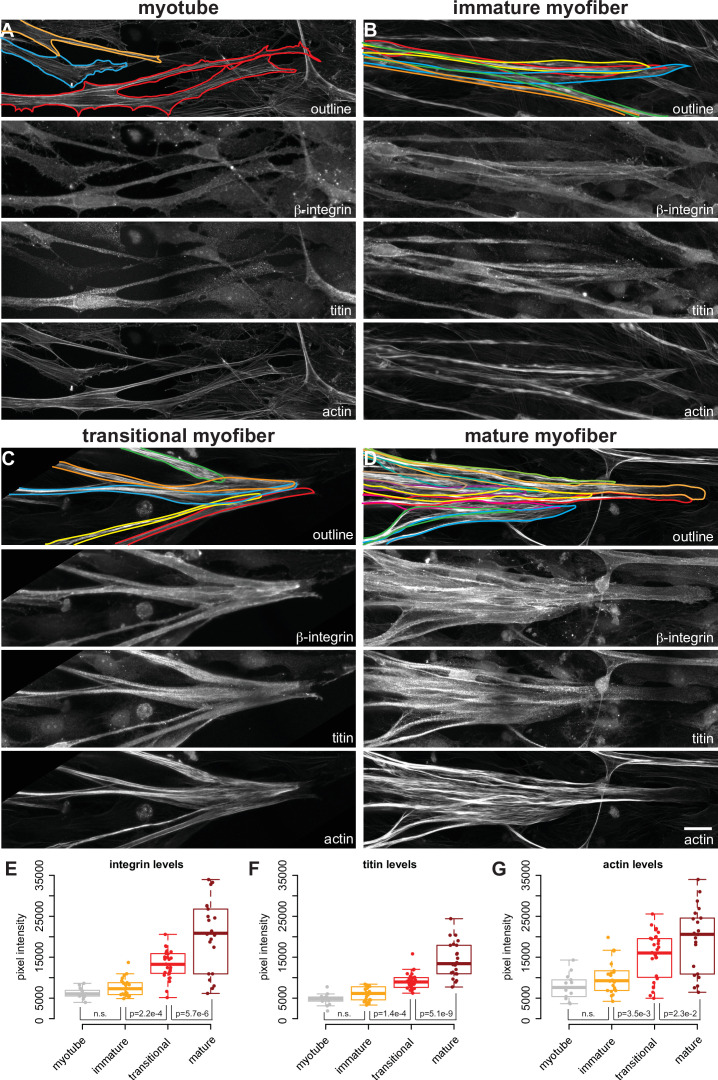
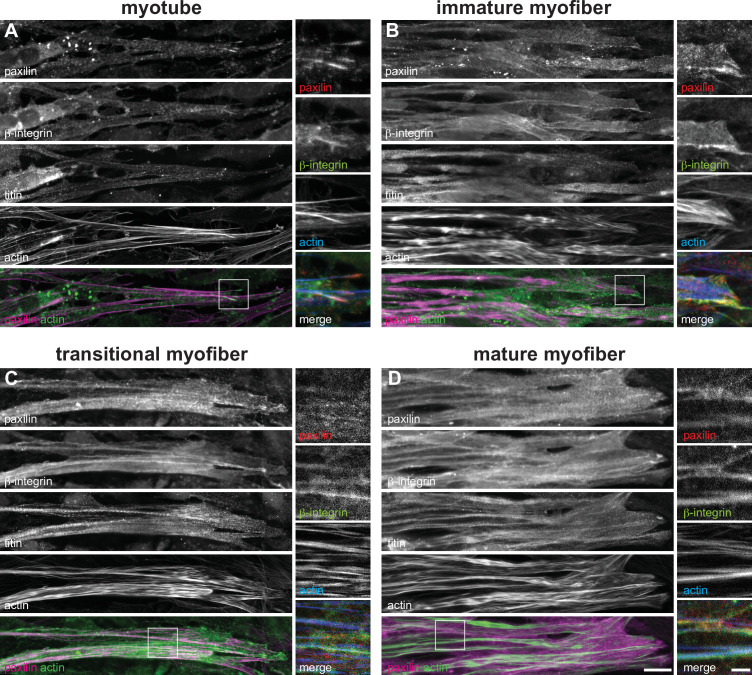
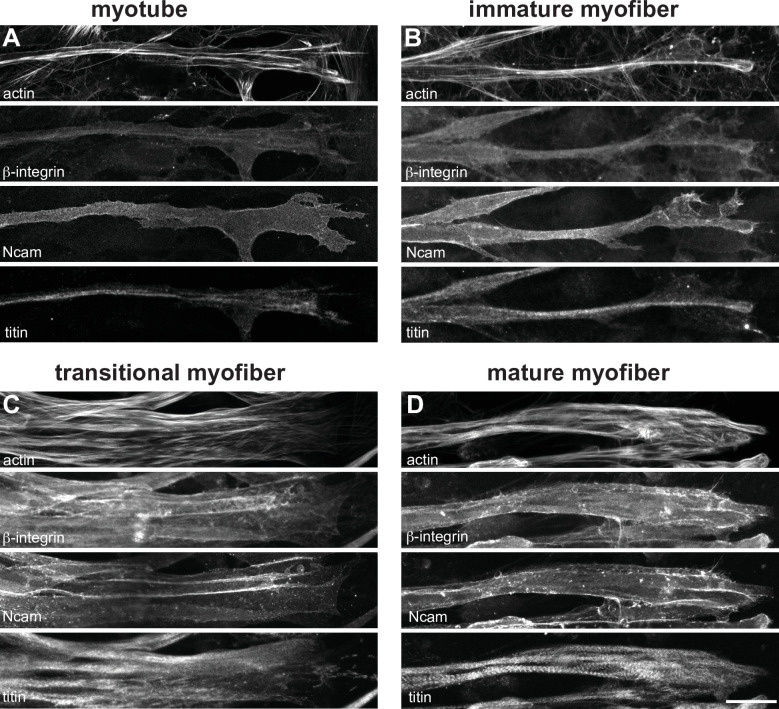
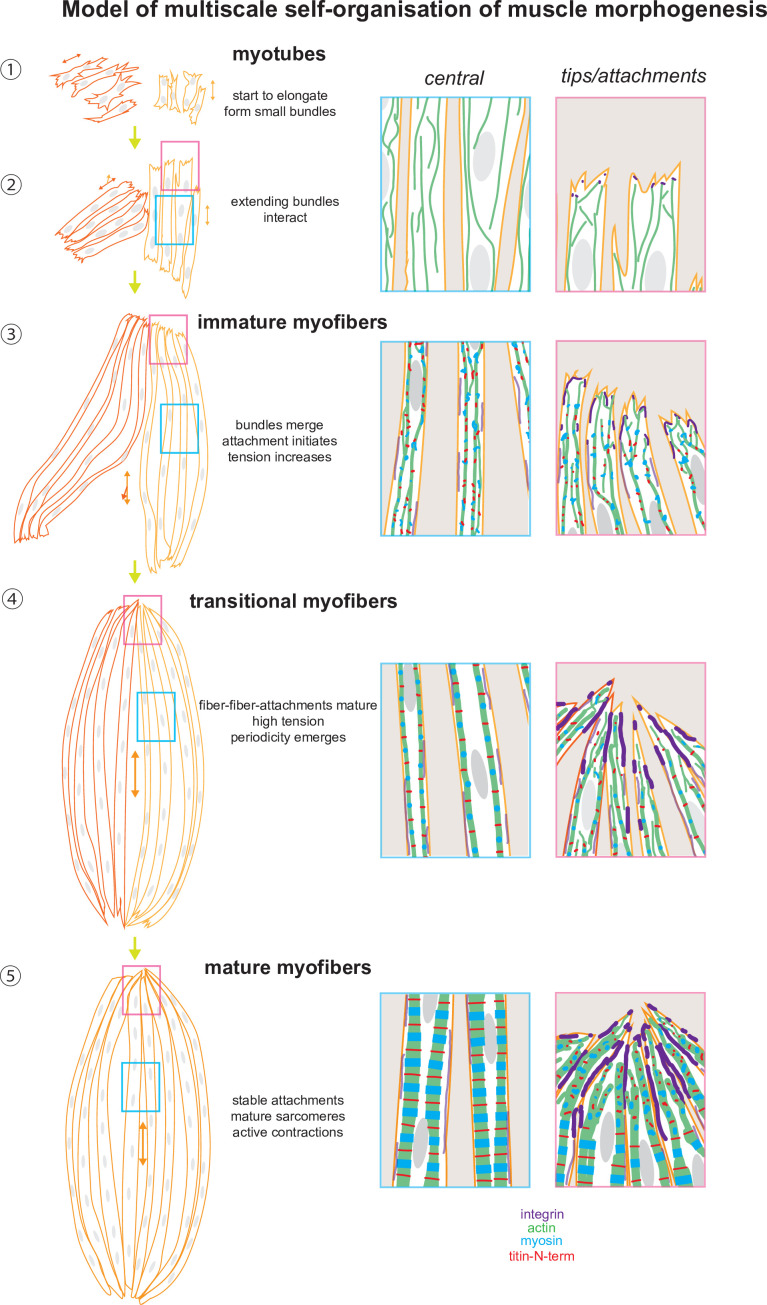
Human muscle is a hierarchically organised tissue with its contractile cells called myofibers packed into large myofiber bundles. Each myofiber contains periodic myofibrils built by hundreds of contractile sarcomeres that generate large mechanical forces. To better understand the mechanisms that coordinate human muscle morphogenesis from tissue to molecular scales, we adopted a simple in vitro system using induced pluripotent stem cell-derived human myogenic precursors. When grown on an unrestricted two-dimensional substrate, developing myofibers spontaneously align and self-organise into higher-order myofiber bundles, which grow and consolidate to stable sizes. Following a transcriptional boost of sarcomeric components, myofibrils assemble into chains of periodic sarcomeres that emerge across the entire myofiber. More efficient myofiber bundling accelerates the speed of sarcomerogenesis suggesting that tension generated by bundling promotes sarcomerogenesis. We tested this hypothesis by directly probing tension and found that tension build-up precedes sarcomere assembly and increases within each assembling myofibril. Furthermore, we found that myofiber ends stably attach to other myofibers using integrin-based attachments and thus myofiber bundling coincides with stable myofiber bundle attachment in vitro. A failure in stable myofiber attachment results in a collapse of the myofibrils. Overall, our results strongly suggest that mechanical tension across sarcomeric components as well as between differentiating myofibers is key to coordinate the multi-scale self-organisation of muscle morphogenesis.
Keywords: developmental biology; human; human induced pluripotent stem cells; mechanical tension; muscle; myofibril; sarcomere; self-organisation.
© 2022, Mao, Acharya et al.
Conflict of interest statement
QM, AA, AR, FM, BD, ZA, JR, MD, AM, EW, CC, VG, PL, BH, OT, OP, FS No competing interests declared
Figures


Similar articles
-
Mechanobiology of muscle and myofibril morphogenesis.Cells Dev. 2021 Dec;168:203760. doi: 10.1016/j.cdev.2021.203760. Epub 2021 Dec 1. Cells Dev. 2021. PMID: 34863916 Review.
-
A computational approach to quantitatively define sarcomere dimensions and arrangement in skeletal muscle.Comput Methods Programs Biomed. 2021 Nov;211:106437. doi: 10.1016/j.cmpb.2021.106437. Epub 2021 Sep 24. Comput Methods Programs Biomed. 2021. PMID: 34624632
-
Mechanical tension and spontaneous muscle twitching precede the formation of cross-striated muscle in vivo.Development. 2017 Apr 1;144(7):1261-1272. doi: 10.1242/dev.140723. Epub 2017 Feb 7. Development. 2017. PMID: 28174246 Free PMC article.
-
A transcriptomics resource reveals a transcriptional transition during ordered sarcomere morphogenesis in flight muscle.Elife. 2018 May 30;7:e34058. doi: 10.7554/eLife.34058. Elife. 2018. PMID: 29846170 Free PMC article.
-
Mechanical forces during muscle development.Mech Dev. 2017 Apr;144(Pt A):92-101. doi: 10.1016/j.mod.2016.11.003. Epub 2016 Nov 29. Mech Dev. 2017. PMID: 27913119 Review.
Cited by
-
Muscle growth by sarcomere divisions.Sci Adv. 2025 Jul 11;11(28):eadw9445. doi: 10.1126/sciadv.adw9445. Epub 2025 Jul 9. Sci Adv. 2025. PMID: 40632866 Free PMC article.
-
Fibroblast growth factor 8b (FGF-8b) enhances myogenesis and inhibits adipogenesis in rotator cuff muscle cell populations in vitro.Proc Natl Acad Sci U S A. 2024 Jan 2;121(1):e2314585121. doi: 10.1073/pnas.2314585121. Epub 2023 Dec 26. Proc Natl Acad Sci U S A. 2024. PMID: 38147545 Free PMC article.
-
Actuating Extracellular Matrices Decouple the Mechanical and Biochemical Effects of Muscle Contraction on Motor Neurons.Adv Healthc Mater. 2025 Mar;14(6):e2403712. doi: 10.1002/adhm.202403712. Epub 2024 Nov 10. Adv Healthc Mater. 2025. PMID: 39523700 Free PMC article.
-
Efficient generation of a self-organizing neuromuscular junction model from human pluripotent stem cells.Nat Commun. 2023 Dec 19;14(1):8043. doi: 10.1038/s41467-023-43781-3. Nat Commun. 2023. PMID: 38114482 Free PMC article.
-
Three-dimensional imaging studies in mice identify cellular dynamics of skeletal muscle regeneration.Dev Cell. 2024 Jun 3;59(11):1457-1474.e5. doi: 10.1016/j.devcel.2024.03.017. Epub 2024 Apr 2. Dev Cell. 2024. PMID: 38569550 Free PMC article.
References
-
- Adler D, Kelly TS. Vioplot: Violin Plot. Version 0.3.6 2021
-
- Al Tanoury Z, Zimmerman JF, Rao J, Sieiro D, McNamara HM, Cherrier T, Rodríguez-delaRosa A, Hick-Colin A, Bousson F, Fugier-Schmucker C, Marchiano F, Habermann B, Chal J, Nesmith AP, Gapon S, Wagner E, Gupta VA, Bassel-Duby R, Olson EN, Cohen AE, Parker KK, Pourquié O. Prednisolone rescues Duchenne muscular dystrophy phenotypes in human pluripotent stem cell-derived skeletal muscle in vitro. PNAS. 2021;118:e2022960118. doi: 10.1073/pnas.2022960118. - DOI - PMC - PubMed
-
- Borchers HW. R package pracma. Version 2.3.3R-Project. 2021 https://cran.r-project.org/web/packages/pracma/pracma.pdf
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
