

Enhanced mitochondrial activity reshapes a gut microbiota profile that delays NASH progression
- PMID: 35921199
- PMCID: PMC10113004
- DOI: 10.1002/hep.32705
Enhanced mitochondrial activity reshapes a gut microbiota profile that delays NASH progression
Abstract
Background and aims: Recent studies suggest that mitochondrial dysfunction promotes progression to NASH by aggravating the gut-liver status. However, the underlying mechanism remains unclear. Herein, we hypothesized that enhanced mitochondrial activity might reshape a specific microbiota signature that, when transferred to germ-free (GF) mice, could delay NASH progression.
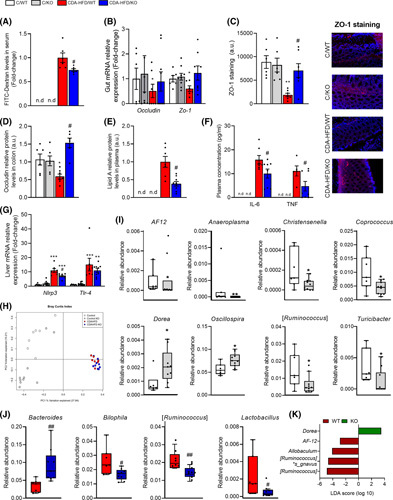
Approach and results: Wild-type and methylation-controlled J protein knockout (MCJ-KO) mice were fed for 6 weeks with either control or a choline-deficient, L-amino acid-defined, high-fat diet (CDA-HFD). One mouse of each group acted as a donor of cecal microbiota to GF mice, who also underwent the CDA-HFD model for 3 weeks. Hepatic injury, intestinal barrier, gut microbiome, and the associated fecal metabolome were then studied. Following 6 weeks of CDA-HFD, the absence of methylation-controlled J protein, an inhibitor of mitochondrial complex I activity, reduced hepatic injury and improved gut-liver axis in an aggressive NASH dietary model. This effect was transferred to GF mice through cecal microbiota transplantation. We suggest that the specific microbiota profile of MCJ-KO, characterized by an increase in the fecal relative abundance of Dorea and Oscillospira genera and a reduction in AF12 , Allboaculum , and [ Ruminococcus ], exerted protective actions through enhancing short-chain fatty acids, nicotinamide adenine dinucleotide (NAD + ) metabolism, and sirtuin activity, subsequently increasing fatty acid oxidation in GF mice. Importantly, we identified Dorea genus as one of the main modulators of this microbiota-dependent protective phenotype.
Conclusions: Overall, we provide evidence for the relevance of mitochondria-microbiota interplay during NASH and that targeting it could be a valuable therapeutic approach.
Copyright © 2023 The Author(s). Published by Wolters Kluwer Health, Inc.
Conflict of interest statement
Dr. Rincón is the Scientific Founder of Mitotherapeutix LLC, and Dr. Martínez‐Chantar advises Mitotherapeutix LLC.
María Juárez-Fernández and Naroa Goikoetxea-Usandizaga joint first authors.
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
