

Accumulation and regulation of anthocyanins in white and purple Tibetan Hulless Barley (Hordeum vulgare L. var. nudum Hook. f.) revealed by combined de novo transcriptomics and metabolomics
- PMID: 35922757
- PMCID: PMC9351122
- DOI: 10.1186/s12870-022-03699-2
Accumulation and regulation of anthocyanins in white and purple Tibetan Hulless Barley (Hordeum vulgare L. var. nudum Hook. f.) revealed by combined de novo transcriptomics and metabolomics
Abstract
Background: Colored barley, which may have associated human health benefits, is more desirable than the standard white variety, but the metabolites and molecular mechanisms underlying seedcoat coloration remain unclear.
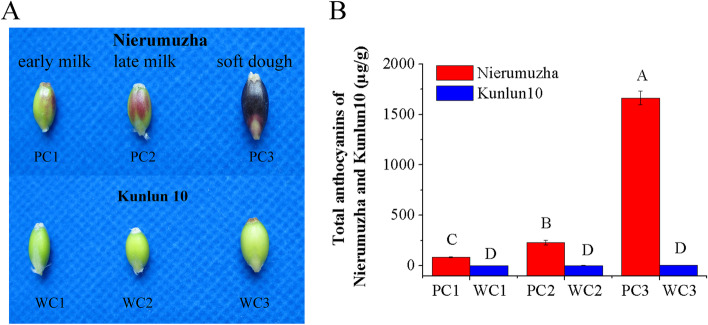
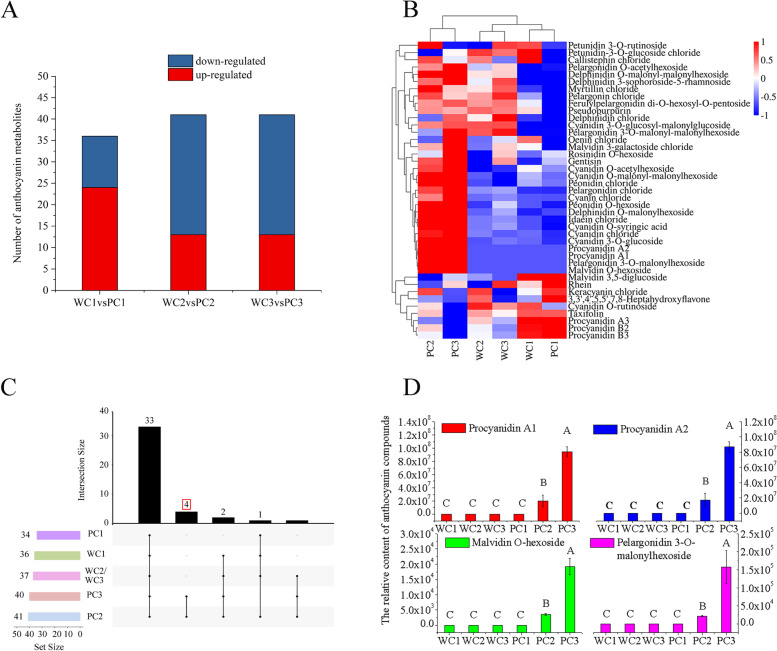
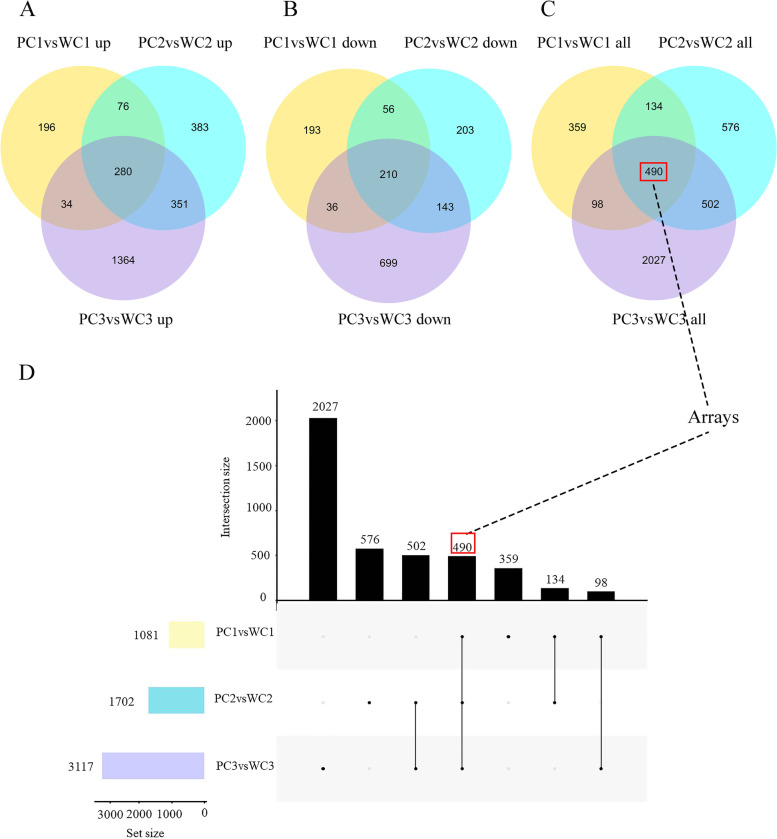
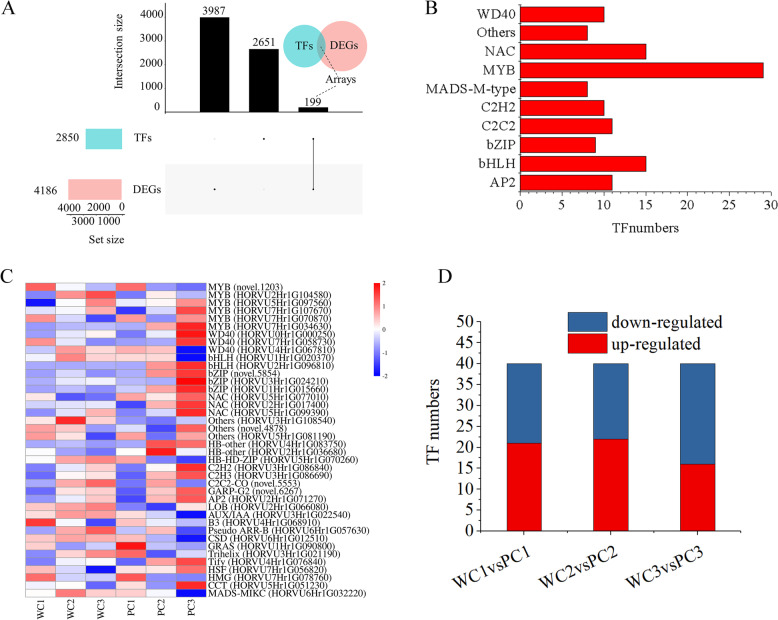
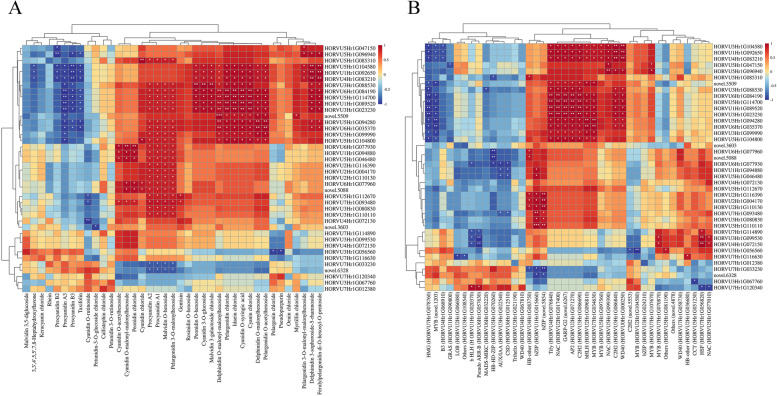
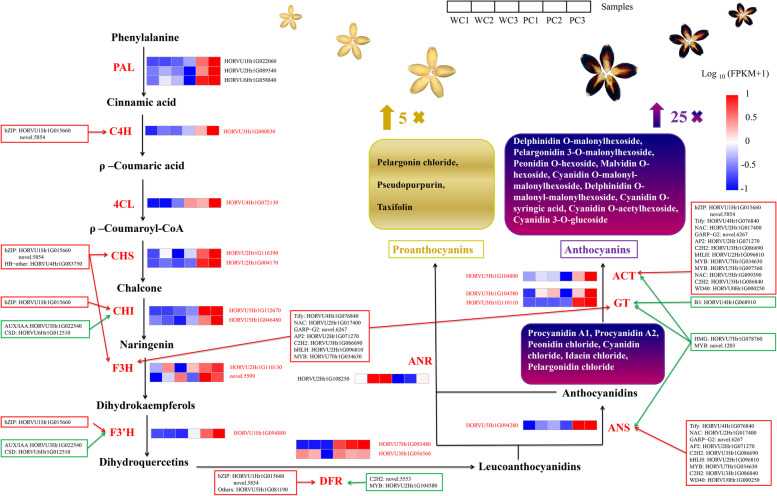
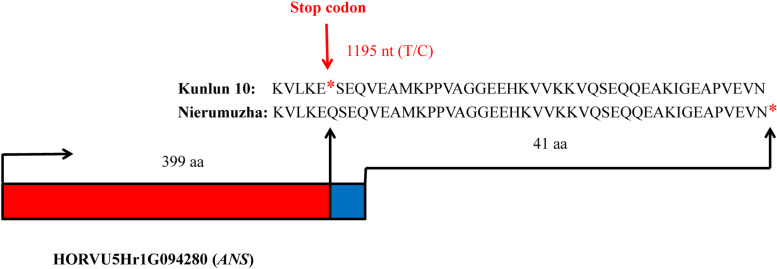
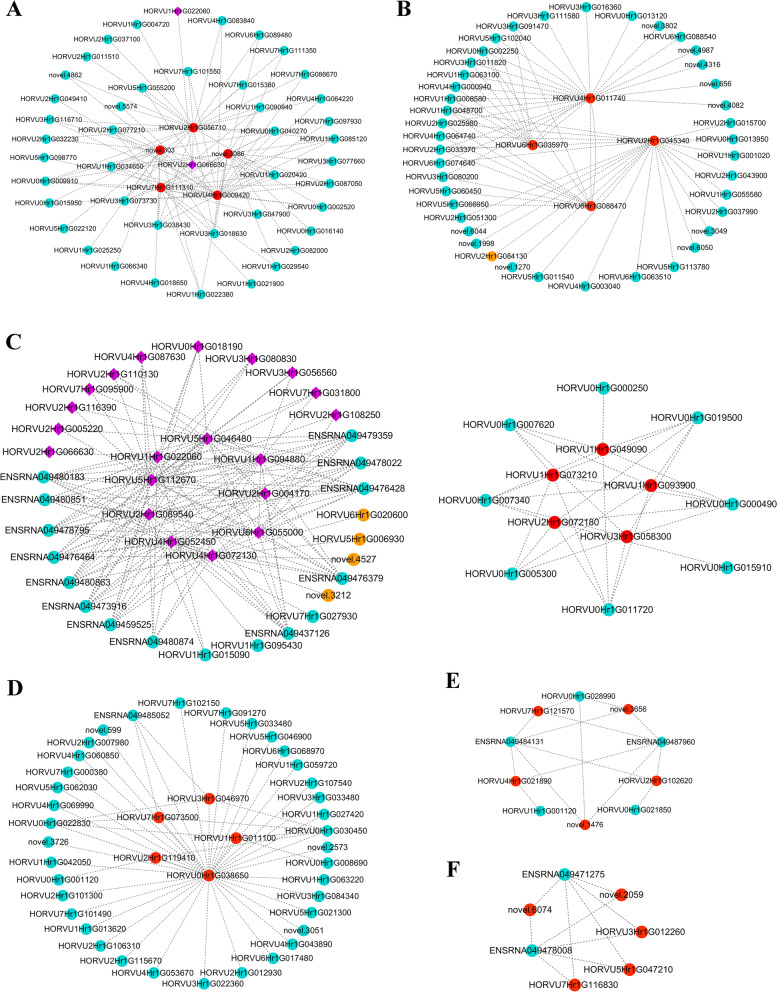
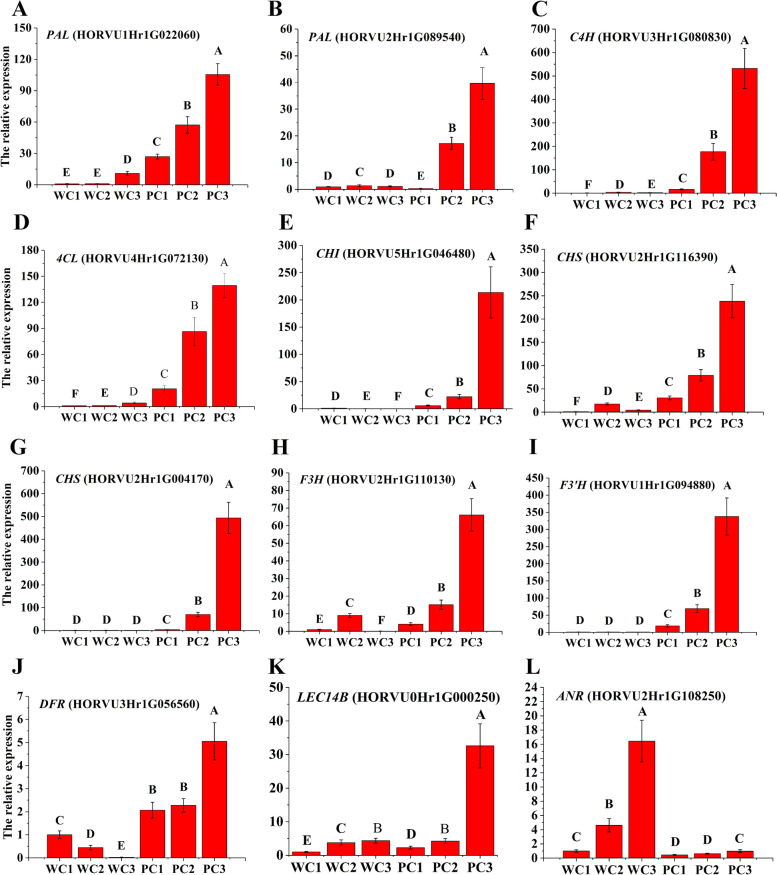
Results: Here, the development of Tibetan hulless barley was monitored, and 18 biological samples at 3 seedcoat color developmental stages were analyzed by transcriptomic and metabolic assays in Nierumuzha (purple) and Kunlun10 (white). A total of 41 anthocyanin compounds and 4186 DEGs were identified. Then we constructed the proanthocyanin-anthocyanin biosynthesis pathway of Tibetan hulless barley, including 19 genes encoding structural enzymes in 12 classes (PAL, C4H, 4CL, CHS, CHI, F3H, F3'H, DFR, ANS, ANR, GT, and ACT). 11 DEGs other than ANR were significantly upregulated in Nierumuzha as compared to Kunlun10, leading to high levels of 15 anthocyanin compounds in this variety (more than 25 times greater than the contents in Kunlun10). ANR was significantly upregulated in Kunlun10 as compared to Nierumuzha, resulting in higher contents of three anthocyanins compounds (more than 5 times greater than the contents in Nierumuzha). In addition, 22 TFs, including MYBs, bHLHs, NACs, bZips, and WD40s, were significantly positively or negatively correlated with the expression patterns of the structural genes. Moreover, comparisons of homologous gene sequences between the two varieties identified 61 putative SNPs in 13 of 19 structural genes. A nonsense mutation was identified in the coding sequence of the ANS gene in Kunlun10. This mutation might encode a nonfunctional protein, further reducing anthocyanin accumulation in Kunlun10. Then we identified 3 modules were highly specific to the Nierumuzha (purple) using WGCNA. Moreover, 12 DEGs appeared both in the putative proanthocyanin-anthocyanin biosynthesis pathway and the protein co-expression network were obtained and verified.
Conclusion: Our study constructed the proanthocyanin-anthocyanin biosynthesis pathway of Tibetan hulless barley. A series of compounds, structural genes and TFs responsible for the differences between purple and white hulless barley were obtained in this pathway. Our study improves the understanding of the molecular mechanisms of anthocyanin accumulation and biosynthesis in barley seeds. It provides new targets for the genetic improvement of anthocyanin content and a framework for improving the nutritional quality of barley.
Keywords: ANS; Proanthocyanin-anthocyanin biosynthesis; Seedcoat color; Tibetan Hulless Barley; Transcriptomic and metabolomic.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Transcriptome Screening of Long Noncoding RNAs and Their Target Protein-Coding Genes Unmasks a Dynamic Portrait of Seed Coat Coloration Associated with Anthocyanins in Tibetan Hulless Barley.Int J Mol Sci. 2023 Jun 24;24(13):10587. doi: 10.3390/ijms241310587. Int J Mol Sci. 2023. PMID: 37445765 Free PMC article.
-
Integrative metabolomic and transcriptomic analyses reveal the mechanisms of Tibetan hulless barley grain coloration.Front Plant Sci. 2022 Oct 25;13:1038625. doi: 10.3389/fpls.2022.1038625. eCollection 2022. Front Plant Sci. 2022. PMID: 36388537 Free PMC article.
-
Purple-grained barley (Hordeum vulgare L.): marker-assisted development of NILs for investigating peculiarities of the anthocyanin biosynthesis regulatory network.BMC Plant Biol. 2019 Feb 15;19(Suppl 1):52. doi: 10.1186/s12870-019-1638-9. BMC Plant Biol. 2019. PMID: 30813902 Free PMC article.
-
Occurrence, Biosynthesis, and Health Benefits of Anthocyanins in Rice and Barley.Int J Mol Sci. 2025 Jun 27;26(13):6225. doi: 10.3390/ijms26136225. Int J Mol Sci. 2025. PMID: 40649998 Free PMC article. Review.
-
Recent insights into anthocyanin biosynthesis, gene involvement, distribution regulation, and domestication process in rice (Oryza sativa L.).Plant Sci. 2024 Dec;349:112282. doi: 10.1016/j.plantsci.2024.112282. Epub 2024 Oct 9. Plant Sci. 2024. PMID: 39389316 Review.
Cited by
-
Investigating the regulatory role of HvANT2 in anthocyanin biosynthesis through protein-motif interaction in Qingke.PeerJ. 2024 Jul 10;12:e17736. doi: 10.7717/peerj.17736. eCollection 2024. PeerJ. 2024. PMID: 39006012 Free PMC article.
-
Comparative Transcriptome and Metabolome Profiling Reveal Mechanisms of Red Leaf Color Fading in Populus × euramericana cv. 'Zhonghuahongye'.Plants (Basel). 2023 Oct 9;12(19):3511. doi: 10.3390/plants12193511. Plants (Basel). 2023. PMID: 37836251 Free PMC article.
-
Genetic analysis and molecular mapping of the purple leaf sheath in barley (Hordeum vulgare).Plant Genome. 2025 Jun;18(2):e70034. doi: 10.1002/tpg2.70034. Plant Genome. 2025. PMID: 40275436 Free PMC article.
-
Potential regulatory genes of light induced anthocyanin accumulation in sweet cherry identified by combining transcriptome and metabolome analysis.Front Plant Sci. 2023 Aug 16;14:1238624. doi: 10.3389/fpls.2023.1238624. eCollection 2023. Front Plant Sci. 2023. PMID: 37662172 Free PMC article.
-
Transcriptome Screening of Long Noncoding RNAs and Their Target Protein-Coding Genes Unmasks a Dynamic Portrait of Seed Coat Coloration Associated with Anthocyanins in Tibetan Hulless Barley.Int J Mol Sci. 2023 Jun 24;24(13):10587. doi: 10.3390/ijms241310587. Int J Mol Sci. 2023. PMID: 37445765 Free PMC article.
References
-
- Yao X, Wang Y, Yao Y, Bai Y, Wu K, Qiao Y. Identification microRNAs and target genes in Tibetan hulless barley to BLS infection. Agron J. 2021;113:2273–2292. doi: 10.1002/agj2.20649. - DOI
-
- Liang J, Deng G, Long H, Pan Z, Wang C, Cai P, et al. Virus-induced silencing of genes encoding LEA protein in Tibetan hulless barley (Hordeum vulgare ssp. vulgare) and their relationship to drought tolerance. Mol Breed. 2012;30:441–451. doi: 10.1007/s11032-011-9633-3. - DOI
-
- Li Y, Long C, Kato K, Yang C, Sato K. Indigenous knowledge and traditional conservation of hulless barley (Hordeum vulgare) germplasm resources in the Tibetan communities of Shangri-la, Yunnan. SW China Genet Resour Crop Evol. 2011;58:645–655. doi: 10.1007/s10722-010-9604-2. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
