

MBL Binding with AhR Controls Th17 Immunity in Silicosis-Associated Lung Inflammation and Fibrosis
- PMID: 35923908
- PMCID: PMC9342710
- DOI: 10.2147/JIR.S357453
MBL Binding with AhR Controls Th17 Immunity in Silicosis-Associated Lung Inflammation and Fibrosis
Abstract
Objective: Mannan-binding lectin (MBL), a soluble pattern recognition molecule of the innate immune system, is primarily synthesized in the liver and secreted into the circulation. Low serum level of MBL has been reported to be related to an increased risk of lung diseases. Herein, we aimed to investigate the function of MBL in silicosis-associated pulmonary inflammation.
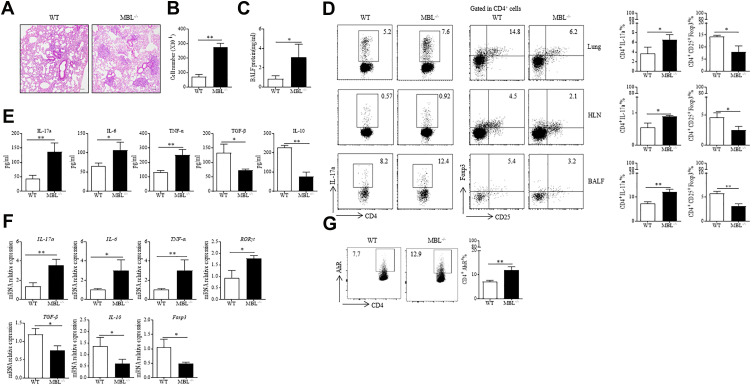
Methods: Serum collected from silicosis patients was tested for correlation between serum MBL levels and Th17 immunity. In vitro studies were performed to further demonstrated the effect of MBL on Th17 polarization. Silica was intratracheally injected in wild type (WT) or MBL-deficient (MBL-/-) mice to induce silicosis-associated lung inflammation and fibrosis. Th17 response was evaluated to explore the effect of MBL on silicosis in vivo.
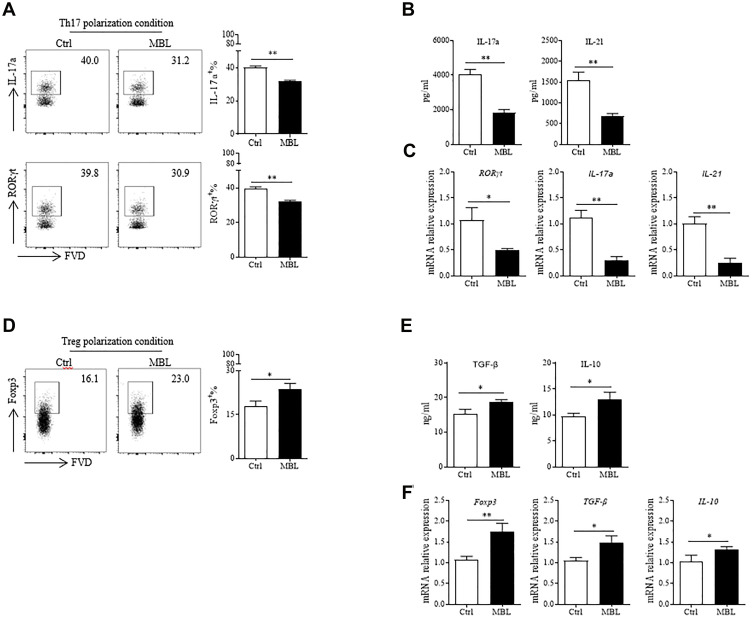
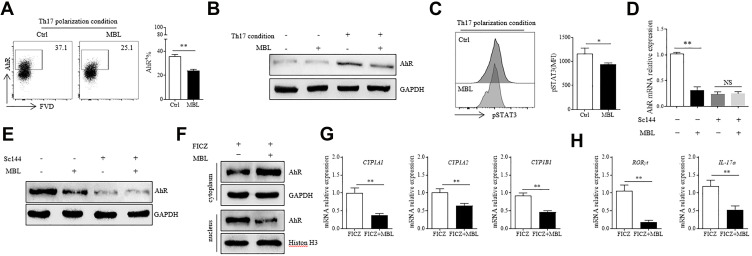
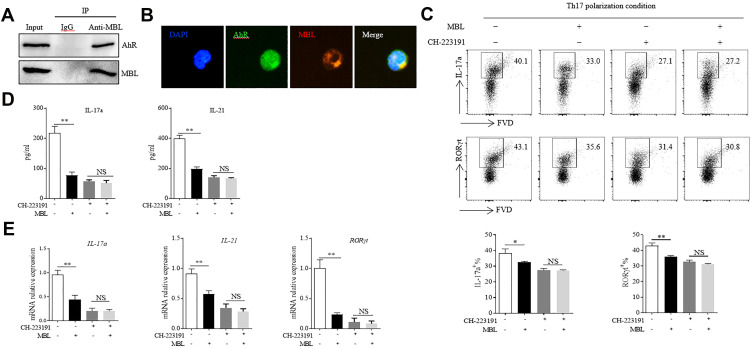
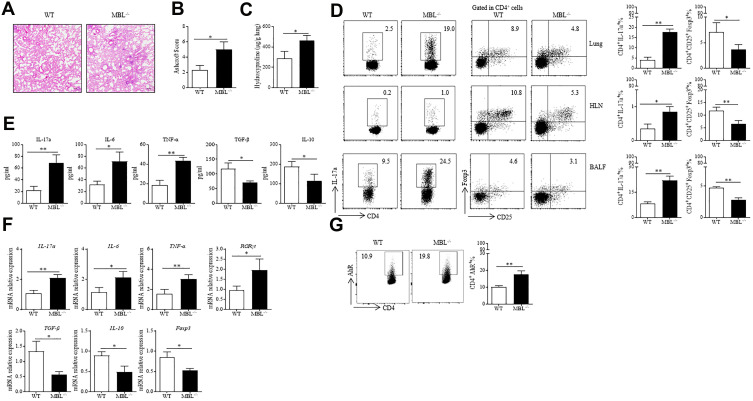
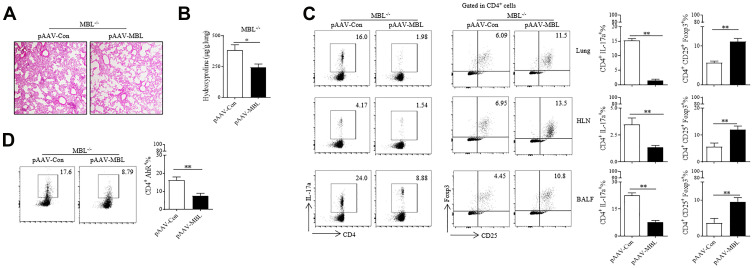
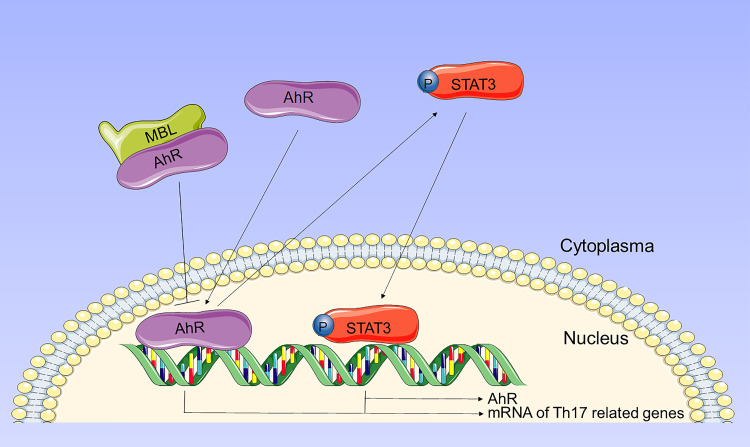
Results: Silicosis patients with high serum MBL levels displayed ameliorative lung function. We demonstrated that serum MBL levels negatively correlated to Th17 cell frequency in silicosis patients. MBL protein markedly reduced expression of IL-17 but enhanced expression of Foxp3 in CD4+ T cells in vitro when subjected to Th17 or Treg polarizing conditions, respectively. The presence of MBL during Th17 cell polarization significantly limited aryl hydrocarbon receptor (AhR) expression and suppressed the signal transducer and activator of transcription 3 (STAT3) phosphorylation. Treatment with the AhR antagonist abolished the effect of MBL on Th17 response. Strikingly, MBL directly bound to AhR and affected its nuclear translocation. Furthermore, MBL-/- mice displayed elevated Th17 cell levels compared with WT mice in response to the silica challenge. The CD4+ T lymphocytes from silica-administrated MBL-/- mice exhibited more AhR expression than the wild-type counterparts.
Conclusion: Our study suggested that MBL limited the Th17 immunity via controlling the AhR/STAT3 pathway, thus providing new insight into silicosis and other inflammatory diseases in patients with MBL deficiency.
Keywords: Th17 cell; aryl hydrocarbon receptor; mannan-binding lectin; signal transducer and activator of transcription 3; silicosis.
© 2022 Liu et al.
Conflict of interest statement
The authors have no conflicts of interest to declare.
Figures

Similar articles
-
Mannan-Binding Lectin Regulates the Th17/Treg Axis Through JAK/STAT and TGF-β/SMAD Signaling Against Candida albicans Infection.J Inflamm Res. 2022 Mar 11;15:1797-1810. doi: 10.2147/JIR.S344489. eCollection 2022. J Inflamm Res. 2022. PMID: 35300210 Free PMC article.
-
Mannan-binding lectin exacerbates the severity of psoriasis by promoting plasmacytoid dendritic cell differentiation via the signal transducer and activator of transcription 3-interferon regulatory factor 8 axis.J Dermatol. 2022 May;49(5):496-507. doi: 10.1111/1346-8138.16323. Epub 2022 Mar 29. J Dermatol. 2022. PMID: 35347767
-
Mannan-binding lectin, a serum collectin, suppresses T-cell proliferation via direct interaction with cell surface calreticulin and inhibition of proximal T-cell receptor signaling.FASEB J. 2017 Jun;31(6):2405-2417. doi: 10.1096/fj.201601200RR. Epub 2017 Feb 16. FASEB J. 2017. PMID: 28209773
-
Tregs promote the differentiation of Th17 cells in silica-induced lung fibrosis in mice.PLoS One. 2012;7(5):e37286. doi: 10.1371/journal.pone.0037286. Epub 2012 May 15. PLoS One. 2012. PMID: 22615967 Free PMC article.
-
New developments in the understanding of immunology in silicosis.Curr Opin Allergy Clin Immunol. 2007 Apr;7(2):168-73. doi: 10.1097/ACI.0b013e32802bf8a5. Curr Opin Allergy Clin Immunol. 2007. PMID: 17351471 Review.
Cited by
-
Lung microbiota: implications and interactions in chronic pulmonary diseases.Front Cell Infect Microbiol. 2024 Aug 19;14:1401448. doi: 10.3389/fcimb.2024.1401448. eCollection 2024. Front Cell Infect Microbiol. 2024. PMID: 39233908 Free PMC article. Review.
-
The Lectin Pathway of the Complement System-Activation, Regulation, Disease Connections and Interplay with Other (Proteolytic) Systems.Int J Mol Sci. 2024 Jan 26;25(3):1566. doi: 10.3390/ijms25031566. Int J Mol Sci. 2024. PMID: 38338844 Free PMC article. Review.
References
-
- Latoche JD, Ufelle AC, Fazzi F, Ganguly K, Leikauf GD, Fattman CL. Secreted phosphoprotein 1 and sex-specific differences in silica-induced pulmonary fibrosis in mice. Environ Health Perspect. 2016;124(8):1199–1207. PubMed PMID: 26955063; PubMed Central PMCID: PMCPMC4977050. doi:10.1289/ehp.1510335 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
