

DKK1 drives immune suppressive phenotypes in intrahepatic cholangiocarcinoma and can be targeted with anti-DKK1 therapeutic DKN-01
- PMID: 35924447
- PMCID: PMC10087034
- DOI: 10.1111/liv.15383
DKK1 drives immune suppressive phenotypes in intrahepatic cholangiocarcinoma and can be targeted with anti-DKK1 therapeutic DKN-01
Abstract
Background and aims: Dickkopf-1 (DKK1) is associated with poor prognosis in intrahepatic cholangiocarcinoma (iCCA), but the mechanisms behind this are unclear. Here, we show that DKK1 plays an immune regulatory role in vivo and inhibition reduces tumour growth.
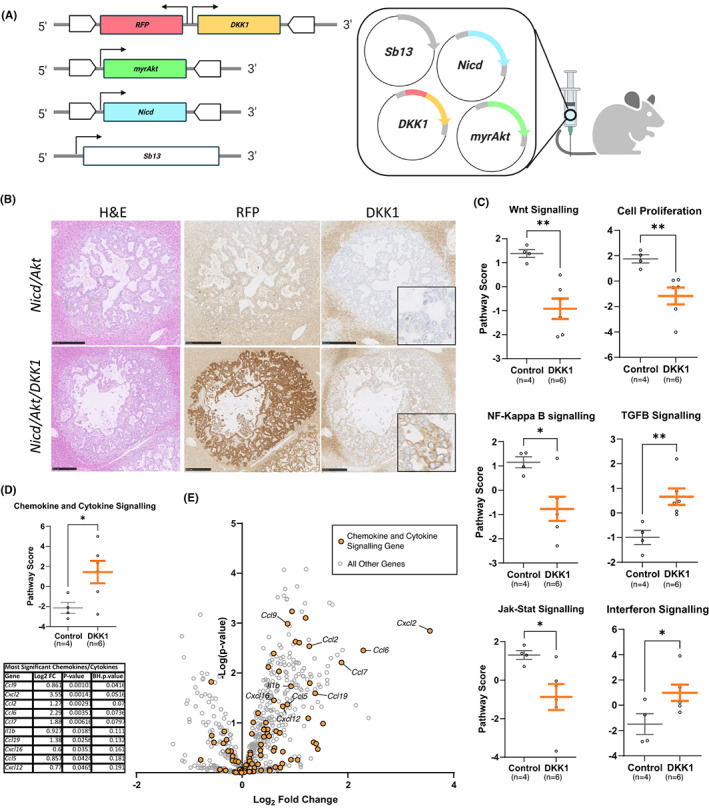
Methods: Various in vivo GEMM mouse models and patient samples were utilized to assess the effects of tumour specific DKK1 overexpression in iCCA. DKK1-driven changes to the tumour immune microenvironment were characterized by immunostaining and gene expression analysis. DKK1 overexpressing and damage-induced models of iCCA were used to demonstrate the therapeutic efficacy of DKK1 inhibition in these contexts using the anti-DKK1 therapeutic, DKN-01.
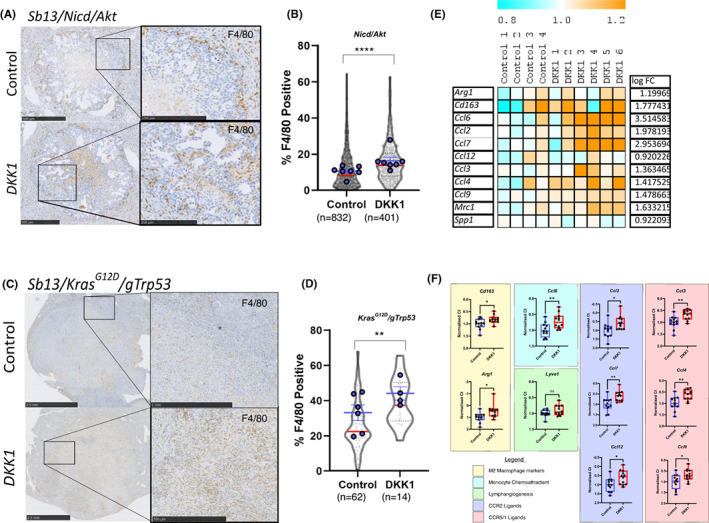
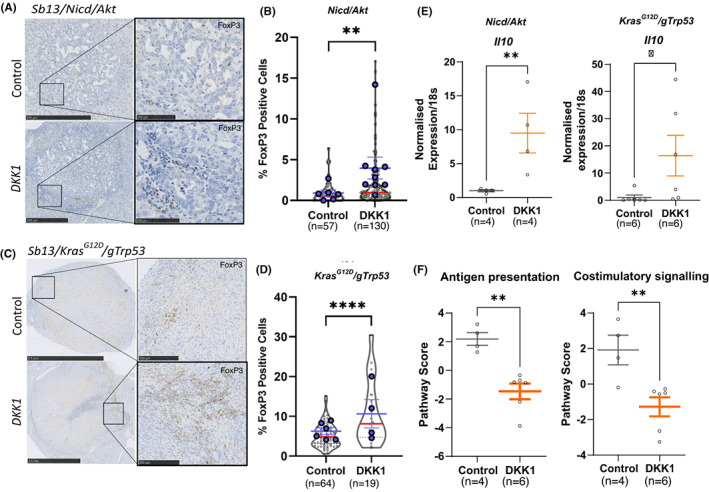
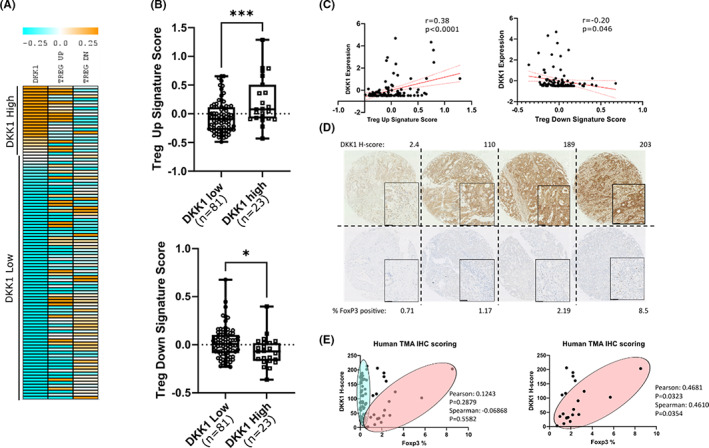
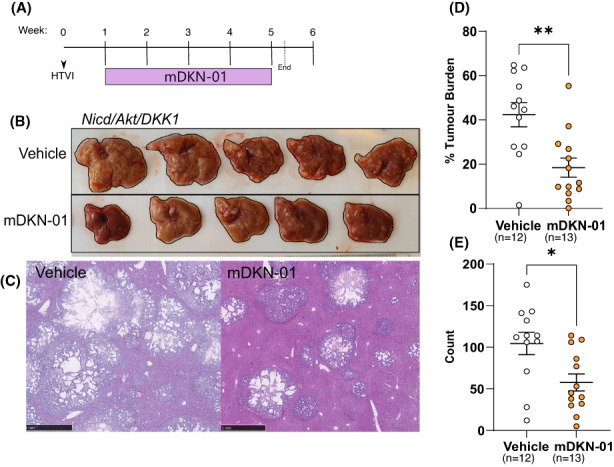
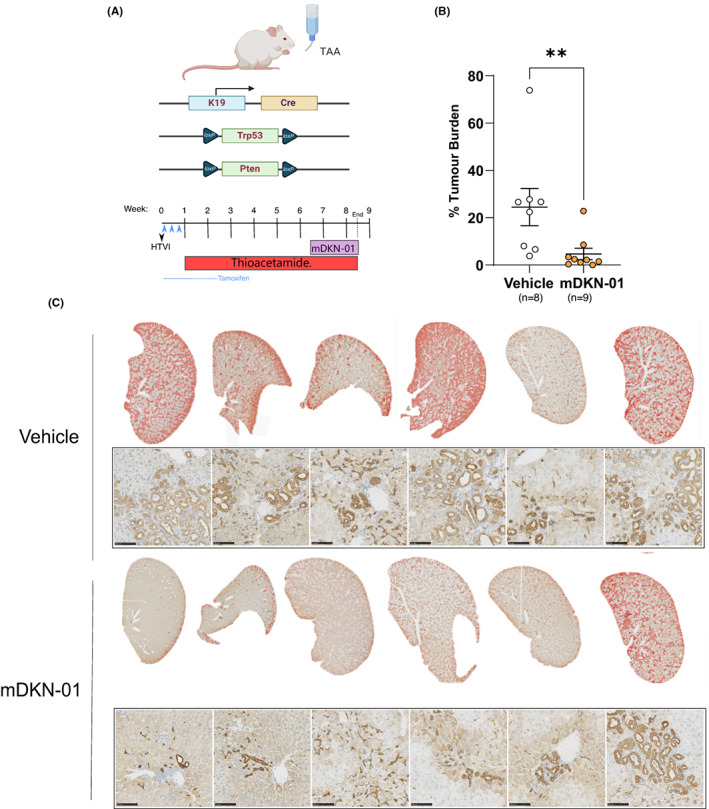
Results: DKK1 overexpression in mouse models of iCCA drives an increase in chemokine and cytokine signalling, the recruitment of regulatory macrophages, and promotes the formation of a tolerogenic niche with higher numbers of regulatory T cells. We show a similar association of DKK1 with FOXP3 and regulatory T cells in patient tissue and gene expression data, demonstrating these effects are relevant to human iCCA. Finally, we demonstrate that inhibition of DKK1 with the monoclonal antibody mDKN-01 is effective at reducing tumour burden in two distinct mouse models of the disease.
Conclusion: DKK1 promotes tumour immune evasion in iCCA through the recruitment of immune suppressive macrophages. Targeting DKK1 with a neutralizing antibody is effective at reducing tumour growth in vivo. As such, DKK1 targeted and immune modulatory therapies may be an effective strategy in iCCA patients with high DKK1 tumour expression or tolerogenic immune phenotypes.
Keywords: Dickkopf-1; cholangiocarcinoma; immune tolerance; macrophage; regulatory T cell.
© 2022 The Authors. Liver International published by John Wiley & Sons Ltd.
Conflict of interest statement
This study was partially funded by Leap Therapeutics. M.K. and W.N. are employees and stockholders and/or stock option holders of Leap Therapeutics Inc. JWP is a founder, shareholder in and on the board of Macomics Ltd an immuno‐oncology company. These studies, however, do not conflict with those of the company. All other authors have no further conflicts to disclose.
Figures
References
-
- Valle JW, Kelley RK, Nervi B, Oh DY, Zhu AX. Biliary tract cancer. Lancet. 2021;397(10272):428‐444. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
