

Genistein Targets STING-Driven Antiviral Responses
- PMID: 35924852
- PMCID: PMC9426420
- DOI: 10.1128/mbio.02064-22
Genistein Targets STING-Driven Antiviral Responses
Abstract
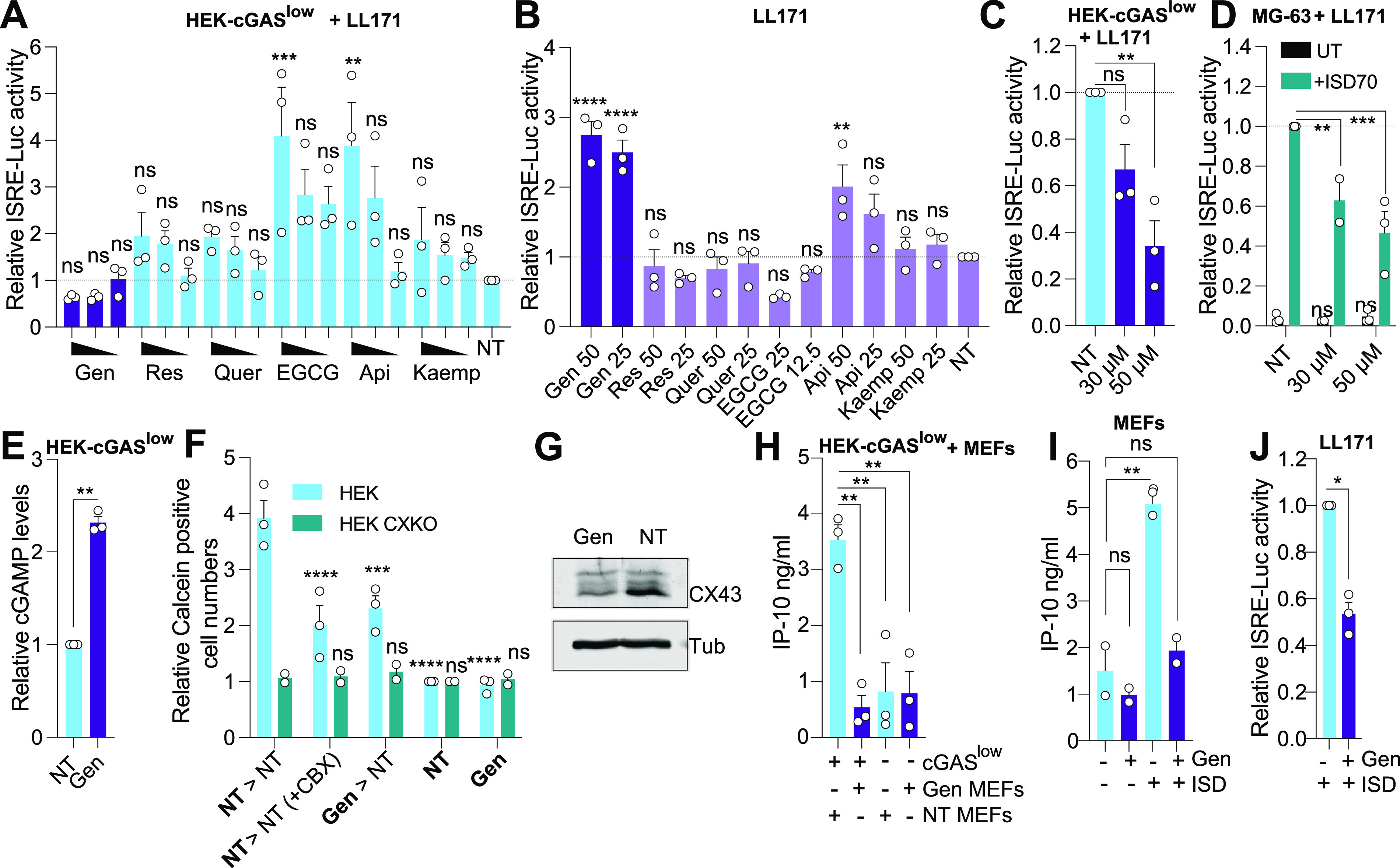
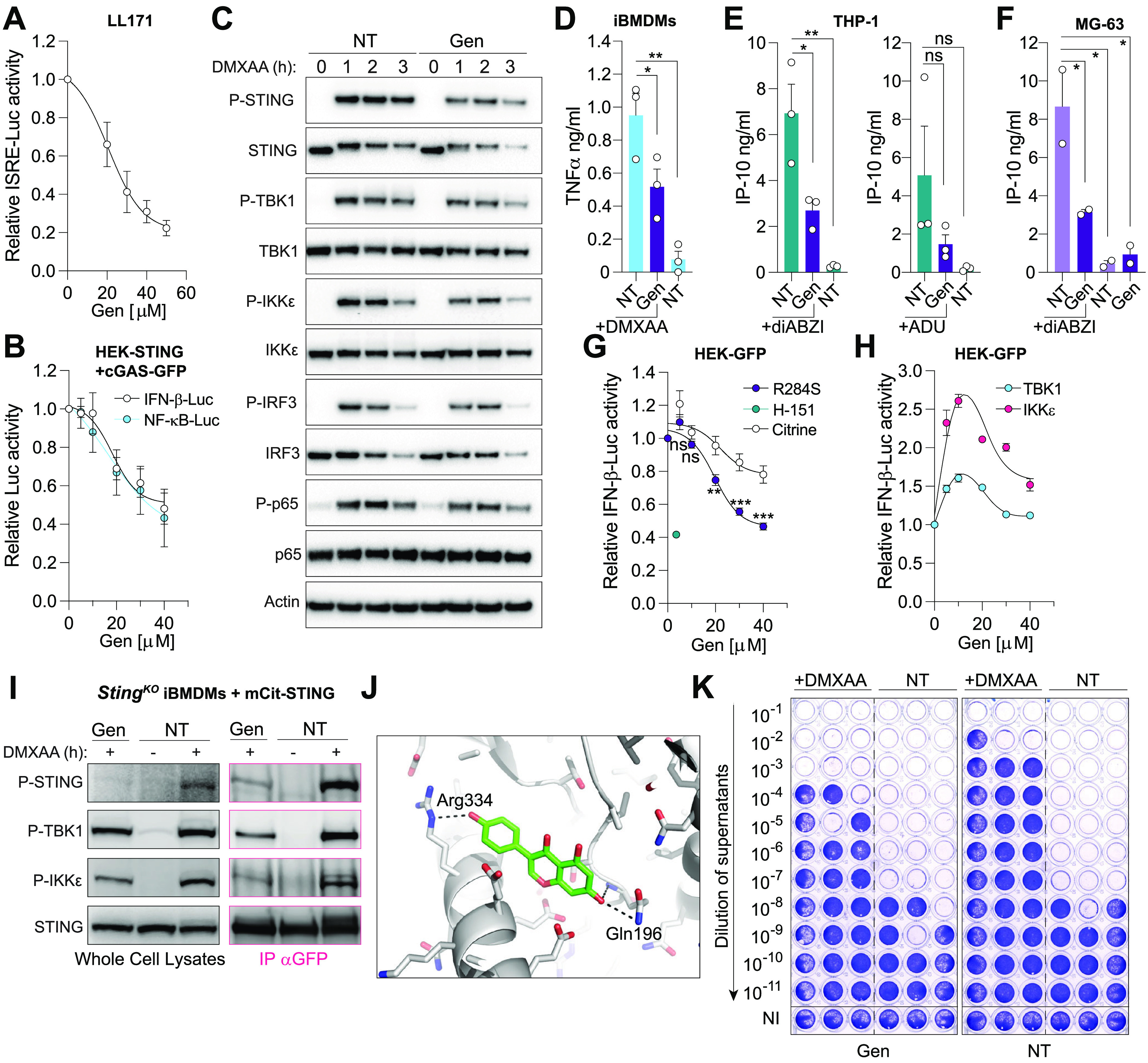
Cytoplasmic detection of DNA by cyclic GMP-AMP (cGAMP) synthase (cGAS) is an essential component of antiviral responses. Upon synthesis, cGAMP binds to the stimulator of interferon (IFN) genes (STING) in infected and adjacent cells through intercellular transfer by connexins forming gap-junctions, eliciting a strong IFN-β-driven antiviral response. We demonstrate here that Genistein, a flavonoid compound naturally occurring in soy-based foods, inhibits cGAS-STING antiviral signaling at two levels. First, Genistein pretreatment of cGAMP-producing cells inhibited gap-junction intercellular communication, resulting in reduced STING responses in adjacent cells. In addition, Genistein directly blocked STING activation by the murine agonist DMXAA, by decreasing the interaction of STING with TBK1 and IKKε. As a result, Genistein attenuated STING signaling in human and mouse cells, dampening antiviral activity against Semliki Forest Virus infection. Collectively, our findings identify a previously unrecognized proviral activity of Genistein mediated via its inhibitory effects at two levels of cGAS-STING signaling. IMPORTANCE Several reports suggest that Genistein exhibits antiviral activities against DNA viruses. Our work uncovers a previously unrecognized proviral effect of Genistein, through inhibition of the cGAS-STING pathway at the level of cGAMP transfer and its sensing by STING. This suggests that the use of Genistein as an antiviral should be taken with caution as it may reduce the protective antiviral effects elicited by host STING activation.
Keywords: Genistein; STING inhibitor; cGAMP; gap junction.
Conflict of interest statement
The authors declare a conflict of interest. Michael Gantier is a named inventor of provisional patents relating to cGAS and STING inhibitors.
Figures
References
-
- Balka KR, Louis C, Saunders TL, Smith AM, Calleja DJ, D'Silva DB, Moghaddas F, Tailler M, Lawlor KE, Zhan Y, Burns CJ, Wicks IP, Miner JJ, Kile BT, Masters SL, De Nardo D. 2020. TBK1 and IKKε act redundantly to mediate STING-induced NF-κB responses in myeloid cells. Cell Rep 31:107492. doi:10.1016/j.celrep.2020.03.056. - DOI - PubMed
-
- Zhao B, Du F, Xu P, Shu C, Sankaran B, Bell SL, Liu M, Lei Y, Gao X, Fu X, Zhu F, Liu Y, Laganowsky A, Zheng X, Ji J-Y, West AP, Watson RO, Li P. 2019. A conserved PLPLRT/SD motif of STING mediates the recruitment and activation of TBK1. Nature 569:718–722. doi:10.1038/s41586-019-1228-x. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
