

Exploiting Violet-Blue Light to Kill Campylobacter jejuni: Analysis of Global Responses, Modeling of Transcription Factor Activities, and Identification of Protein Targets
- PMID: 35924857
- PMCID: PMC9426514
- DOI: 10.1128/msystems.00454-22
Exploiting Violet-Blue Light to Kill Campylobacter jejuni: Analysis of Global Responses, Modeling of Transcription Factor Activities, and Identification of Protein Targets
Abstract
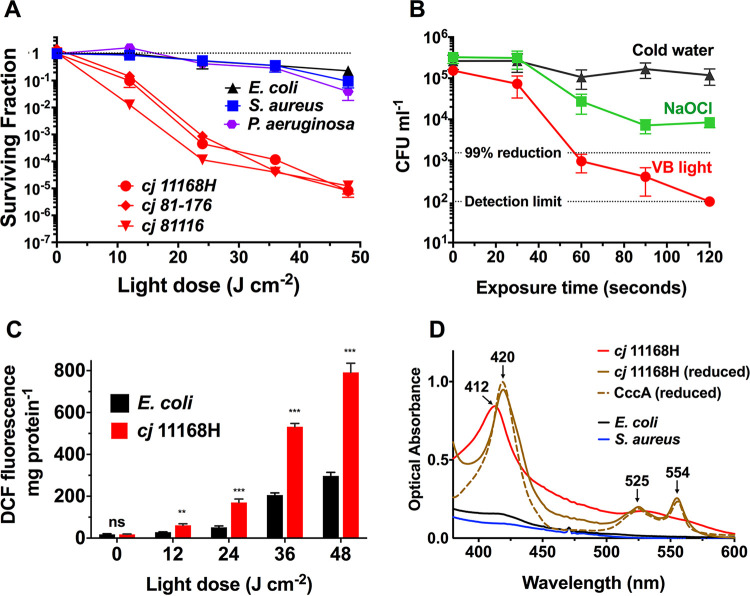
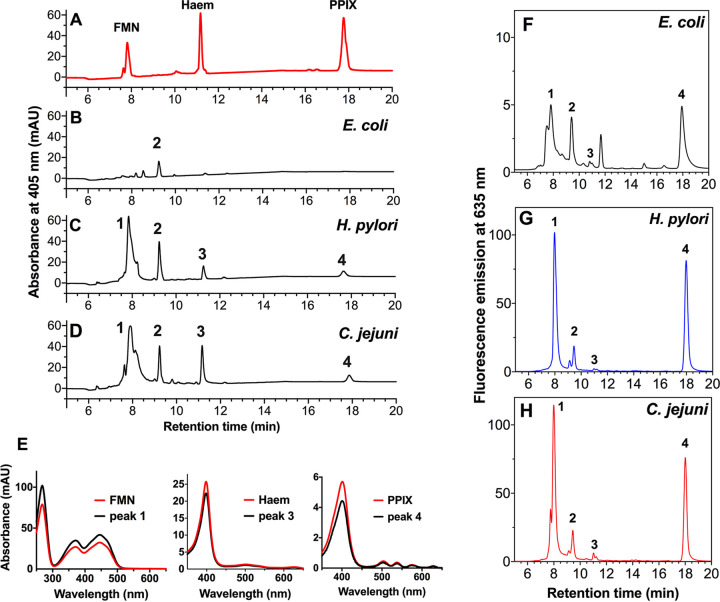
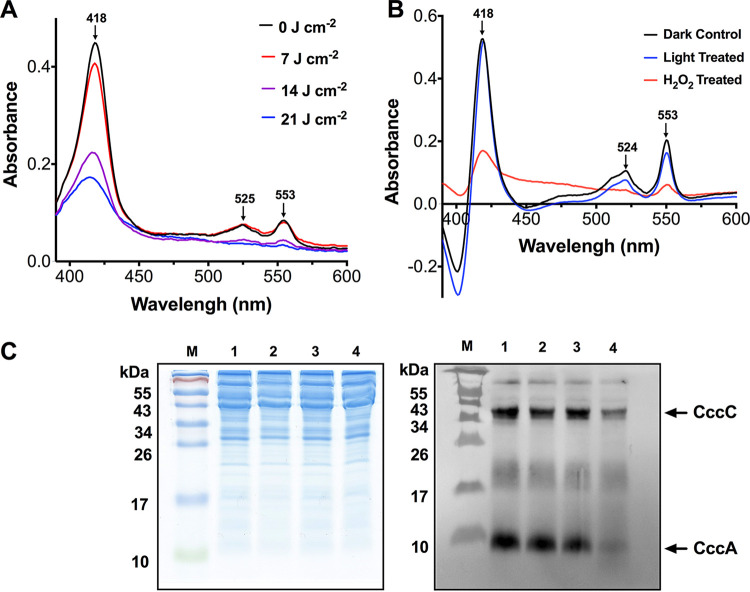
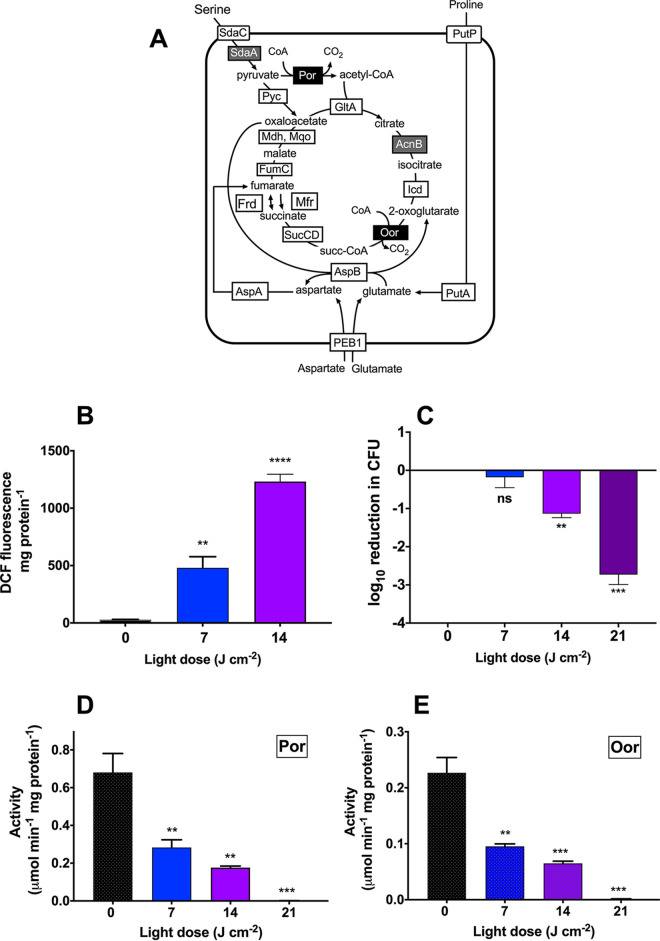
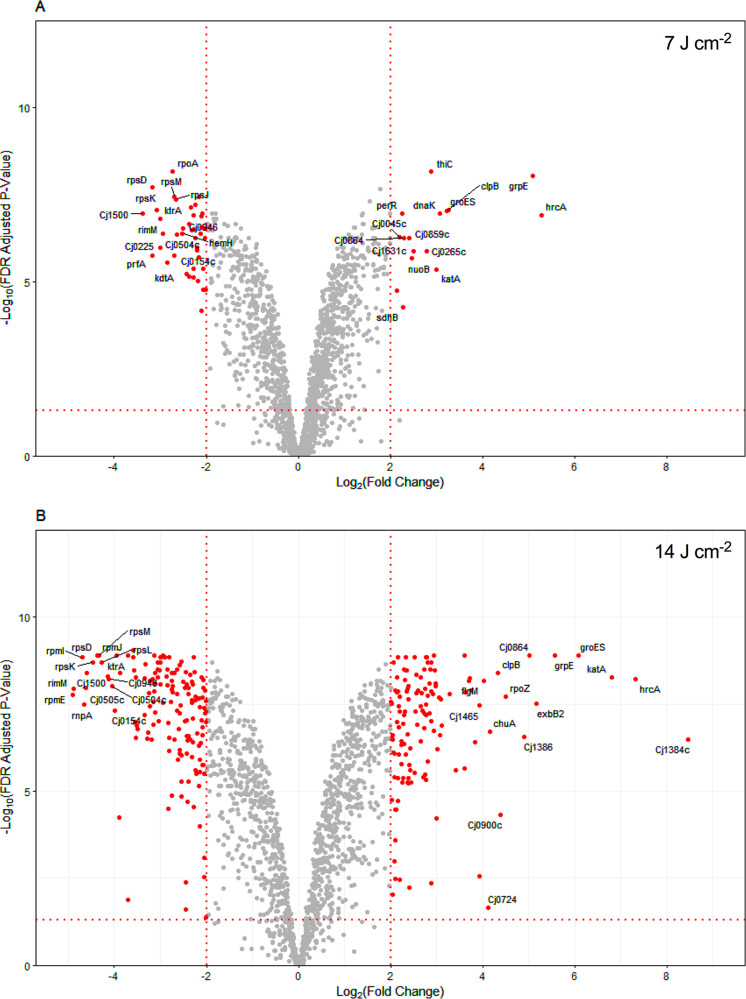
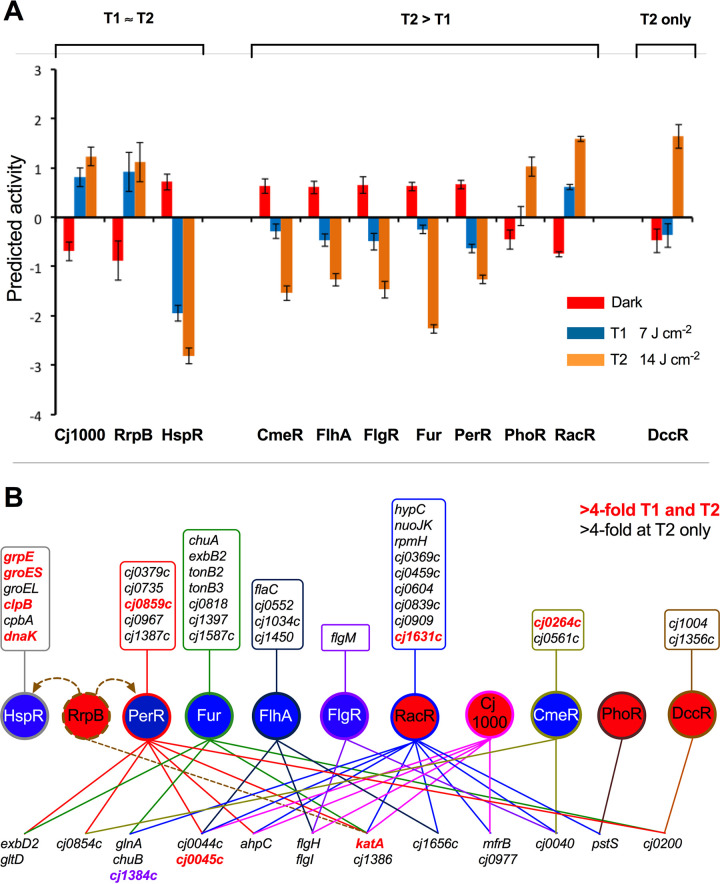
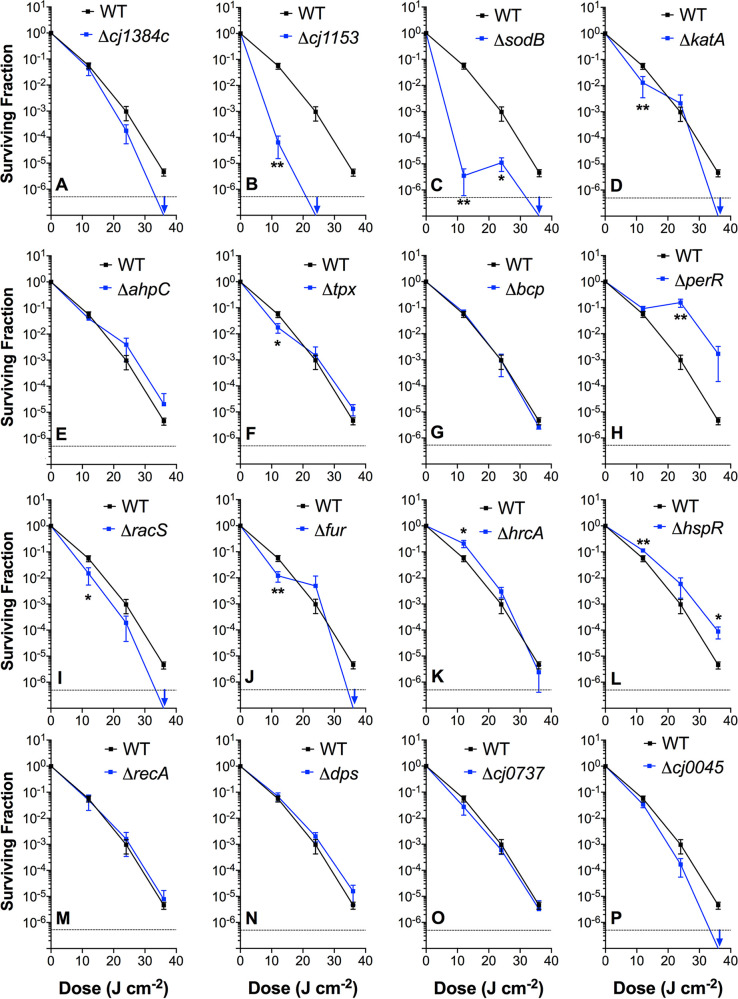
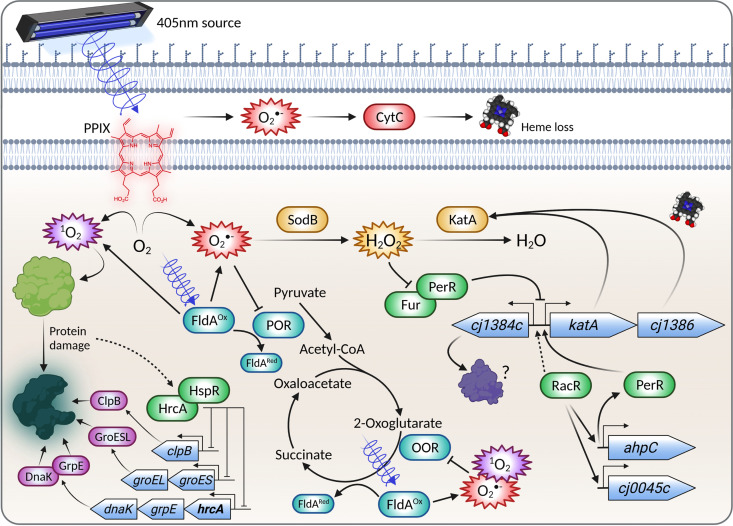
Campylobacter jejuni is a microaerophilic foodborne zoonotic pathogen of worldwide concern as the leading cause of bacterial gastroenteritis. Many strains are increasingly antibiotic resistant and new methods of control are required to reduce food-chain contamination. One possibility is photodynamic inactivation (PDI) using violet-blue (VB) light, to which C. jejuni is highly susceptible. Here, we show that flavin and protoporphyrin IX are major endogenous photosensitizers and that exposure of cells to VB light increases intracellular reactive oxygen species (ROS) to high levels, as indicated by a dichlorodihydrofluorescein reporter. Unusually for an oxygen-respiring bacterium, C. jejuni employs several ROS-sensitive iron-sulfur cluster enzymes in central metabolic pathways; we show that VB light causes rapid inactivation of both pyruvate and 2-oxoglutarate oxidoreductases, thus interrupting the citric acid cycle. Cells exposed to VB light also lose heme from c-type cytochromes, restricting electron transport, likely due to irreversible oxidation of heme-ligating cysteine residues. Evaluation of global gene expression changes by RNAseq and probabilistic modeling showed a two-stage protein damage/oxidative stress response to VB light, driven by specific regulators, including HspR, PerR, Fur, and RacR. Deletion mutant analysis showed that superoxide dismutase and the cytochrome CccA were particularly important for VB light survival and that abolishing repression of chaperones and oxidative stress resistance genes by HcrA, HspR, or PerR increased tolerance to VB light. Our results explain the high innate sensitivity of C. jejuni to VB light and provide new insights that may be helpful in exploiting PDI for novel food-chain interventions to control this pathogen. IMPORTANCE Campylobacteriosis caused by C. jejuni is one of the most widespread zoonotic enteric diseases worldwide and represents an enormous human health and economic burden, compounded by the emergence of antibiotic-resistant strains. New interventions are urgently needed to reduce food-chain contamination. Although UV light is well known to be bactericidal, it is highly mutagenic and problematic for continuous exposure in food production facilities; in contrast, narrow spectrum violet-blue (VB) light is much safer. We confirmed that C. jejuni is highly susceptible to VB light and then identified some of the global regulatory networks involved in responding to photo-oxidative damage. The identification of damaged cellular components underpins efforts to develop commercial applications of VB light-based technologies.
Keywords: RNAseq; flavin; microaerophile; oxidative stress; photodynamic inactivation; protoporphyrin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
PerR controls oxidative stress defence and aerotolerance but not motility-associated phenotypes of Campylobacter jejuni.Microbiology (Reading). 2015 Jul;161(7):1524-36. doi: 10.1099/mic.0.000109. Epub 2015 May 12. Microbiology (Reading). 2015. PMID: 25968890
-
Campylobacter jejuni contains two fur homologs: characterization of iron-responsive regulation of peroxide stress defense genes by the PerR repressor.J Bacteriol. 1999 Oct;181(20):6371-6. doi: 10.1128/JB.181.20.6371-6376.1999. J Bacteriol. 1999. PMID: 10515927 Free PMC article.
-
Characterization of the oxidative stress stimulon and PerR regulon of Campylobacter jejuni.BMC Genomics. 2009 Oct 18;10:481. doi: 10.1186/1471-2164-10-481. BMC Genomics. 2009. PMID: 19835633 Free PMC article.
-
Regulation of oxidative stress resistance in Campylobacter jejuni, a microaerophilic foodborne pathogen.Front Microbiol. 2015 Jul 29;6:751. doi: 10.3389/fmicb.2015.00751. eCollection 2015. Front Microbiol. 2015. PMID: 26284041 Free PMC article. Review.
-
The function, biogenesis and regulation of the electron transport chains in Campylobacter jejuni: New insights into the bioenergetics of a major food-borne pathogen.Adv Microb Physiol. 2019;74:239-329. doi: 10.1016/bs.ampbs.2019.02.003. Epub 2019 Mar 8. Adv Microb Physiol. 2019. PMID: 31126532 Review.
Cited by
-
Efficacy of Violet-Blue (405 nm) LED Lamps for Disinfection of High-Environmental-Contact Surfaces in Healthcare Facilities: Leading to the Inactivation of Microorganisms and Reduction of MRSA Contamination.Pathogens. 2023 Nov 10;12(11):1338. doi: 10.3390/pathogens12111338. Pathogens. 2023. PMID: 38003802 Free PMC article.
-
Blue Light Compromises Bacterial β-Lactamases Activity to Overcome β-Lactam Resistance.Lasers Surg Med. 2024 Sep;56(7):673-681. doi: 10.1002/lsm.23819. Epub 2024 Jul 22. Lasers Surg Med. 2024. PMID: 39039622
-
Identification and validation of reference genes for quantitative gene expression analysis under 409 and 415 nm antimicrobial blue light treatment.Front Mol Biosci. 2025 Jan 6;11:1467726. doi: 10.3389/fmolb.2024.1467726. eCollection 2024. Front Mol Biosci. 2025. PMID: 39834786 Free PMC article.
-
Temporal dynamics of gene expression during the development of Campylobacter jejuni biofilms.Microb Genom. 2025 May;11(5):001387. doi: 10.1099/mgen.0.001387. Microb Genom. 2025. PMID: 40327030 Free PMC article.
-
New Insights into the Bacterial Targets of Antimicrobial Blue Light.Microbiol Spectr. 2023 Feb 21;11(2):e0283322. doi: 10.1128/spectrum.02833-22. Online ahead of print. Microbiol Spectr. 2023. PMID: 36809152 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
