

METTL14-dependent m6A modification controls iNKT cell development and function
- PMID: 35926466
- PMCID: PMC9495716
- DOI: 10.1016/j.celrep.2022.111156
METTL14-dependent m6A modification controls iNKT cell development and function
Abstract
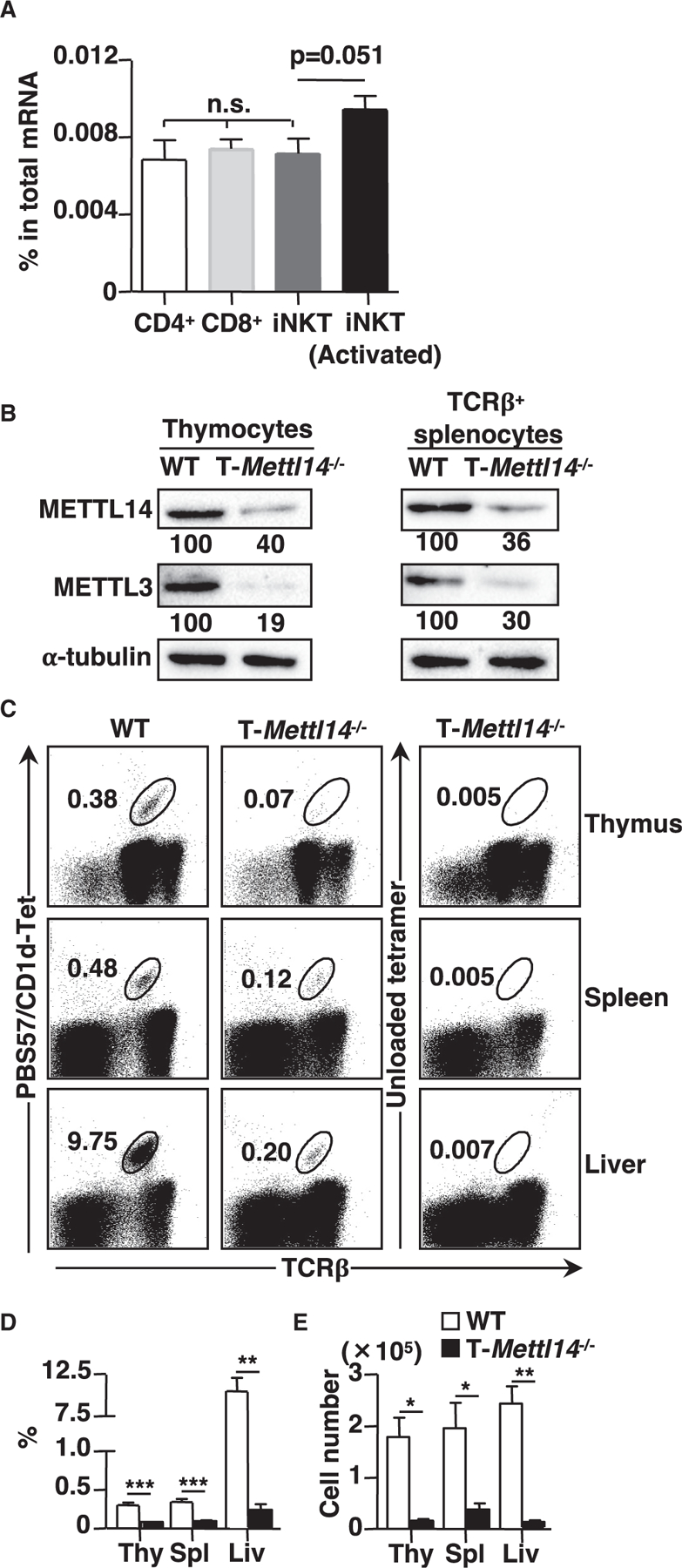
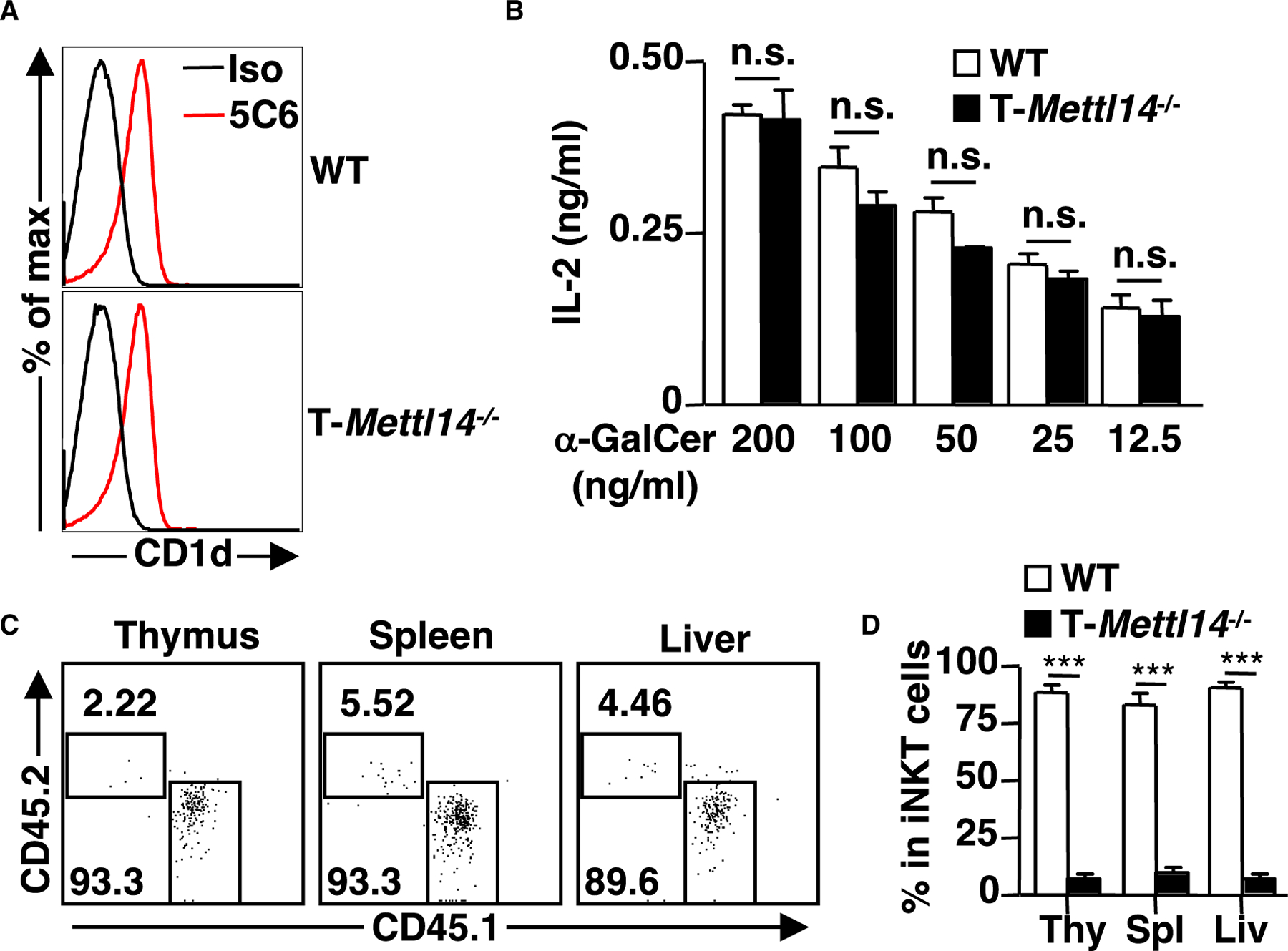
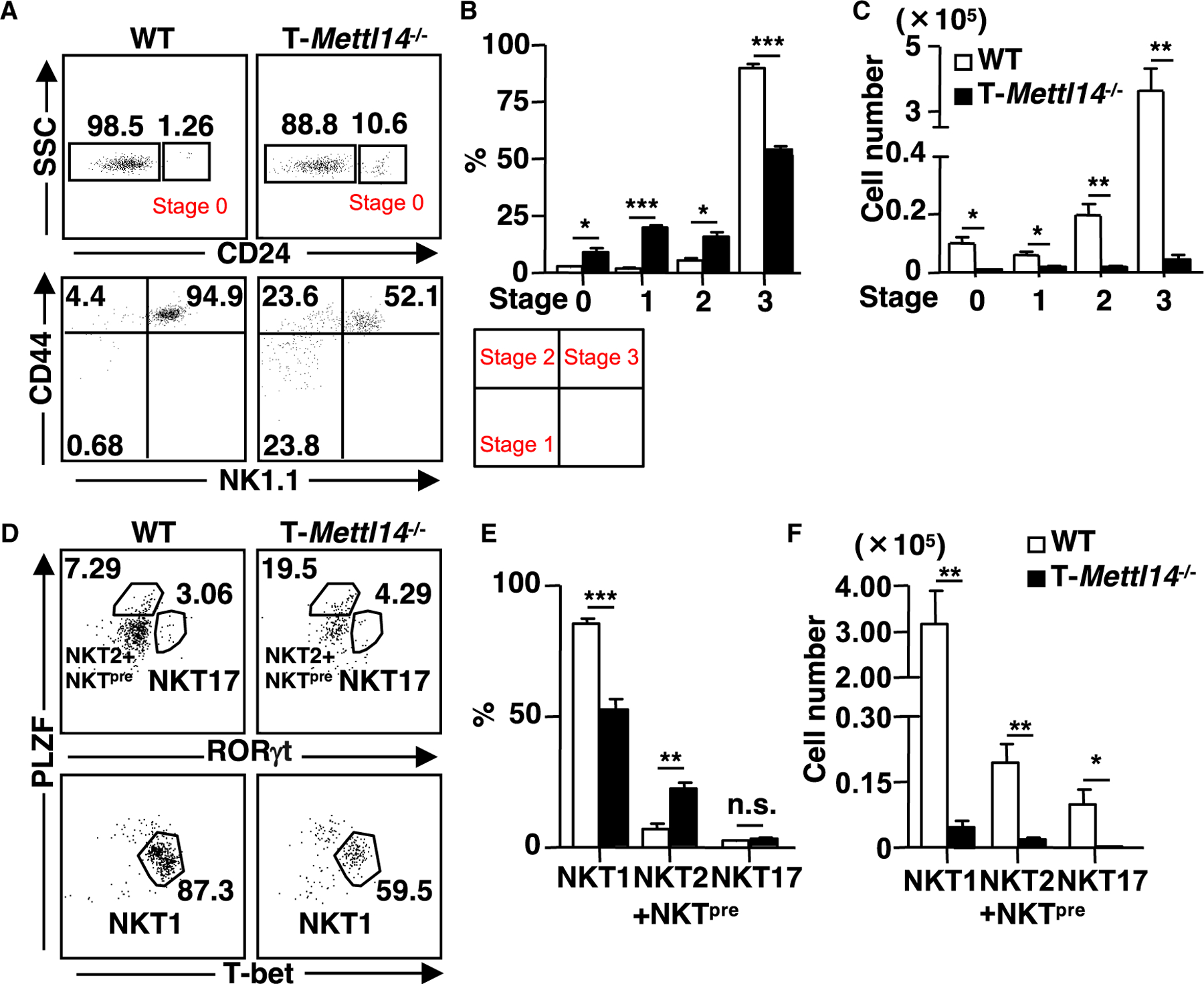
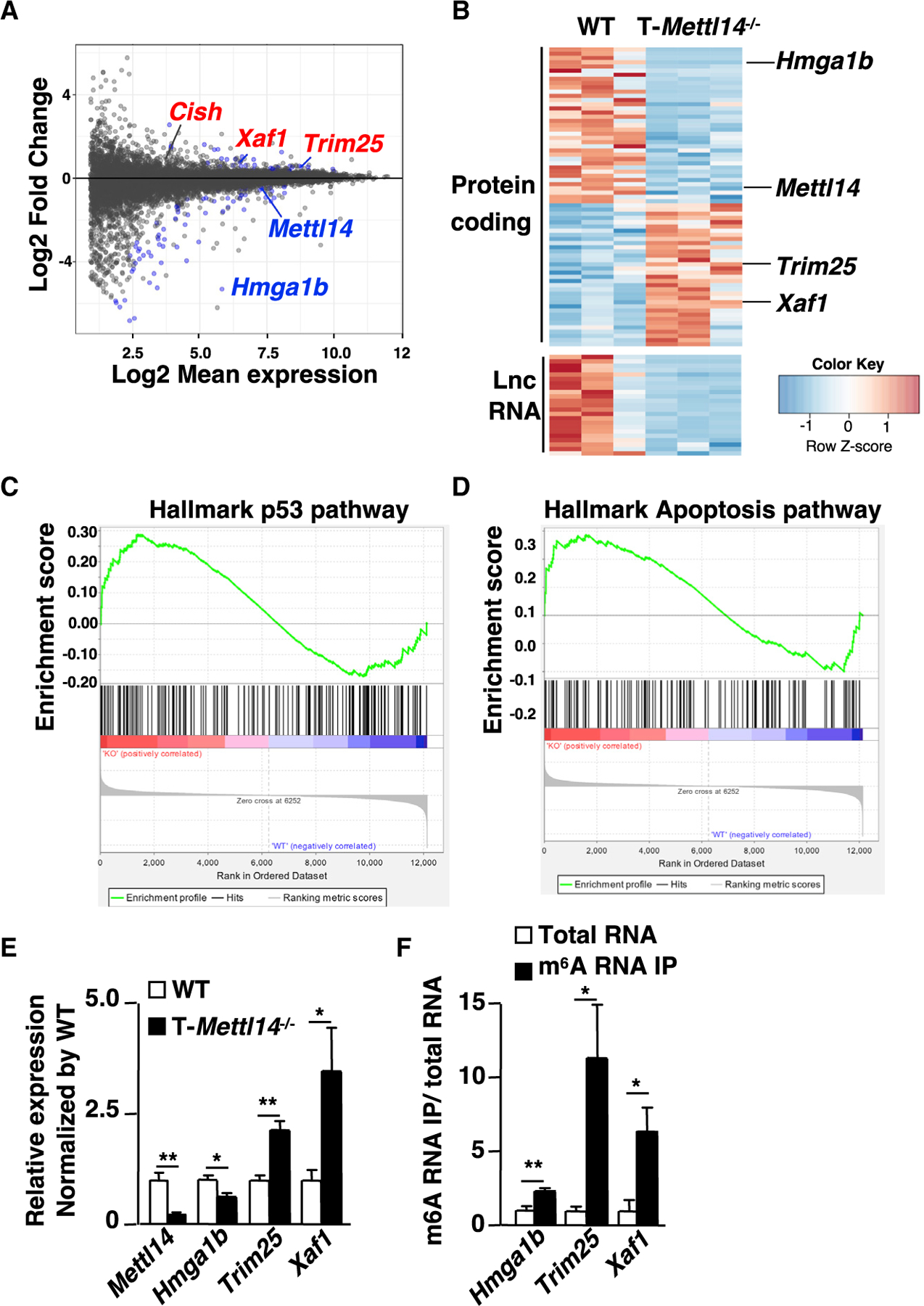
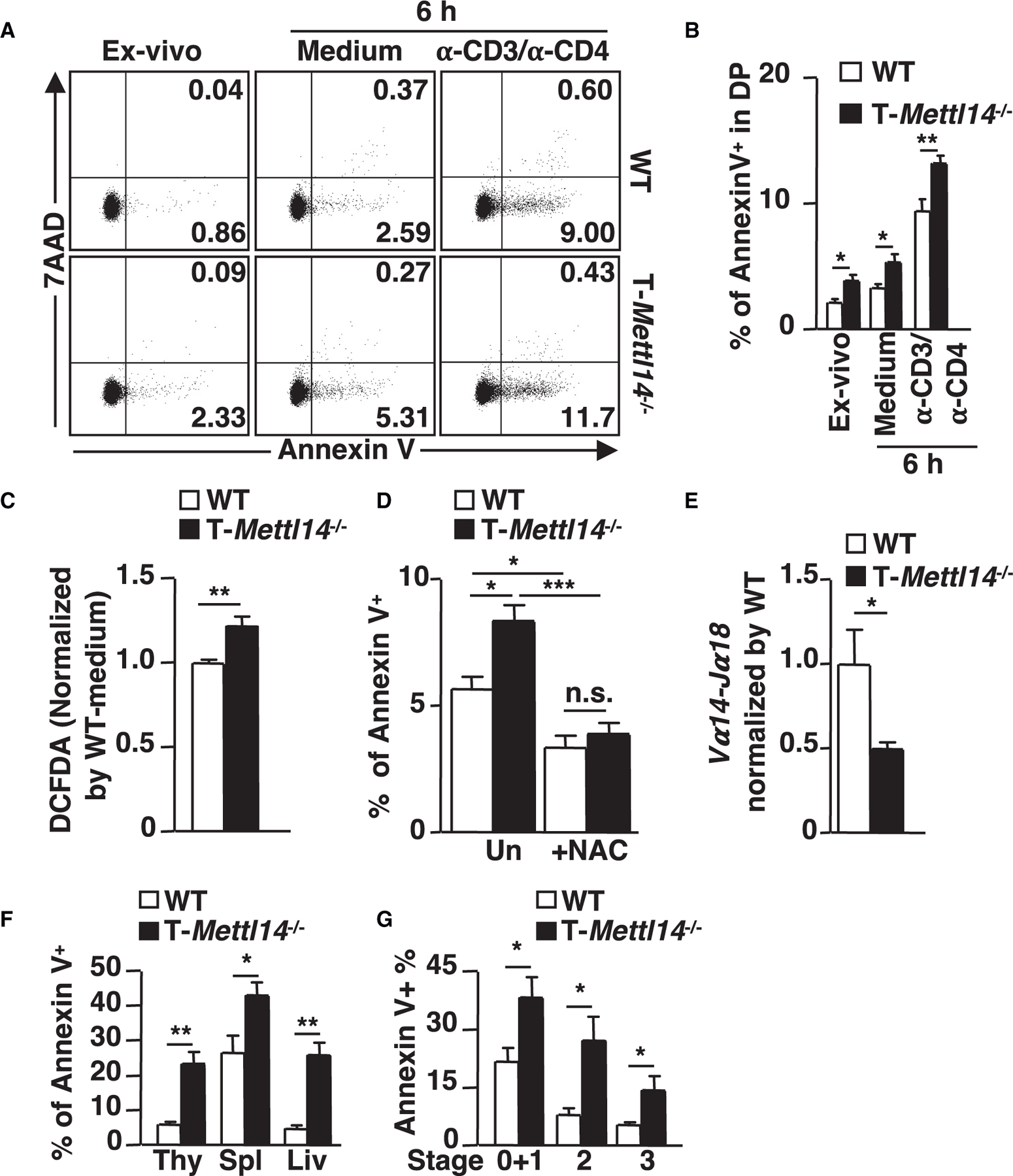
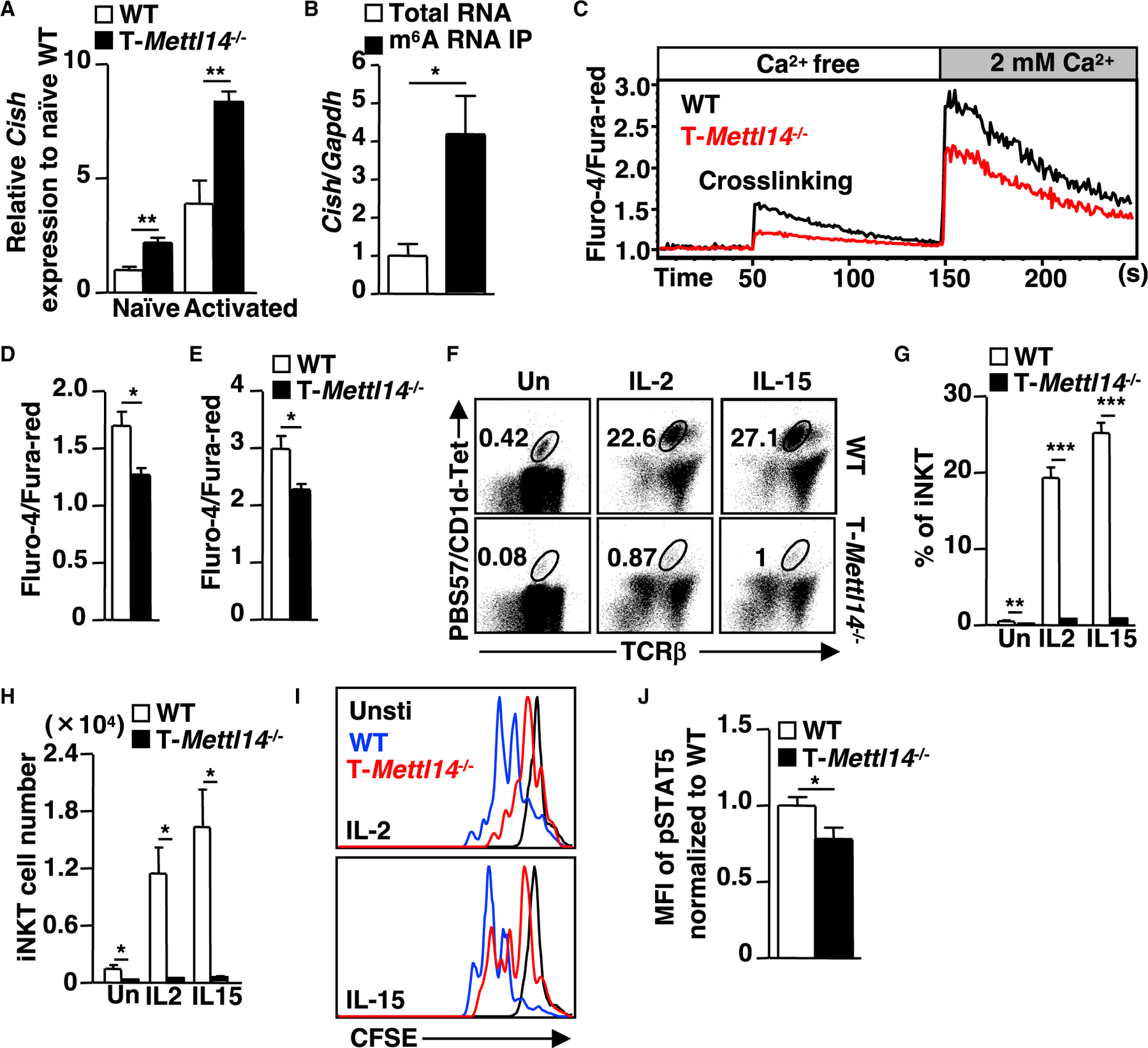
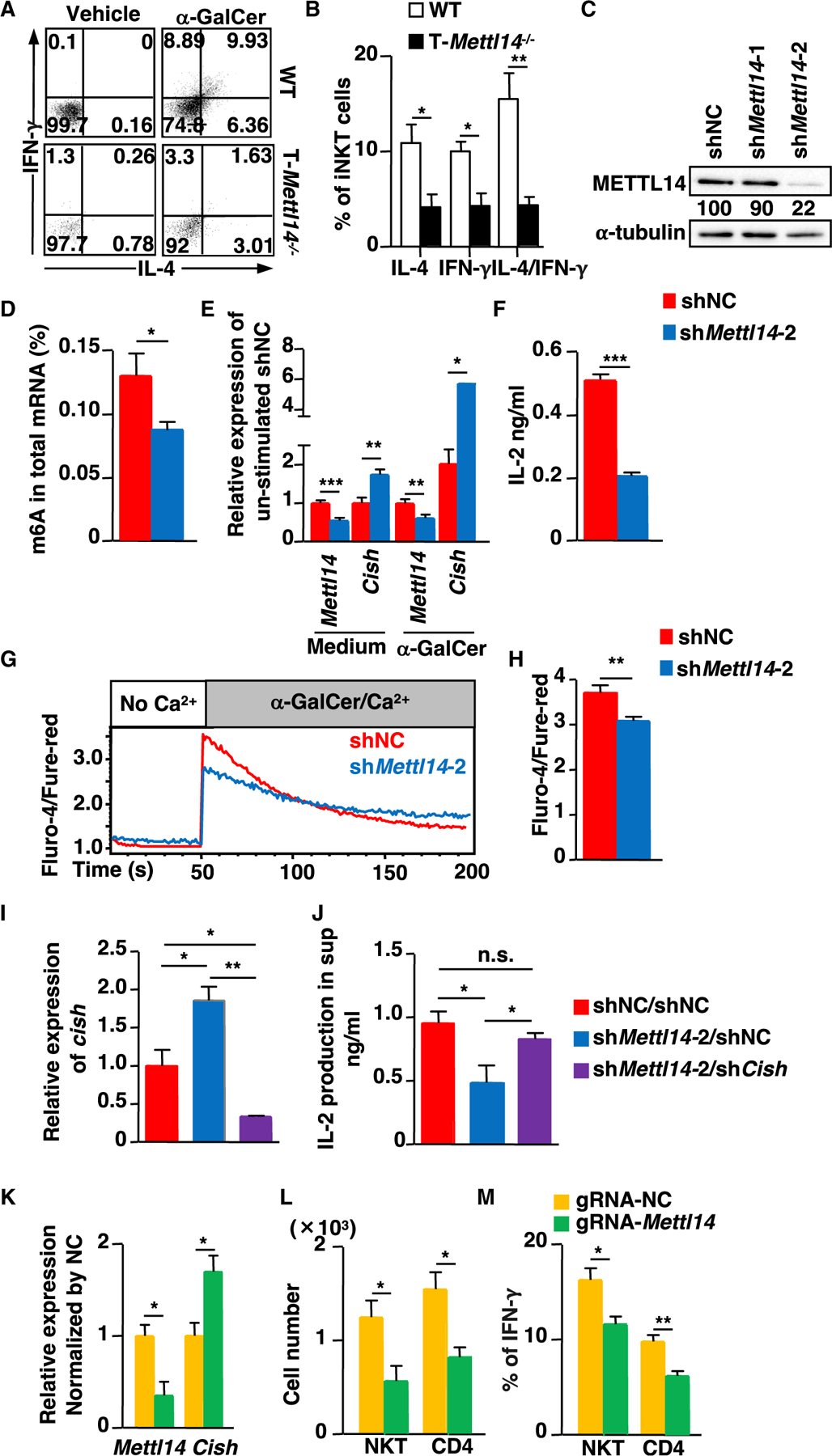
N6-methyladenosine (m6A), the most common form of RNA modification, controls CD4+ T cell homeostasis by targeting the IL-7/STAT5/SOCS signaling pathways. The role of m6A modification in unconventional T cell development remains unknown. Using mice with T cell-specific deletion of RNA methyltransferase METTL14 (T-Mettl14-/-), we demonstrate that m6A modification is indispensable for iNKT cell homeostasis. Loss of METTL14-dependent m6A modification leads to the upregulation of apoptosis in double-positive thymocytes, which in turn decreases Vα14-Jα18 gene rearrangements, resulting in drastic reduction of iNKT numbers in the thymus and periphery. Residual T-Mettl14-/- iNKT cells exhibit increased apoptosis, impaired maturation, and decreased responsiveness to IL-2/IL-15 and TCR stimulation. Furthermore, METTL14 knockdown in mature iNKT cells diminishes their cytokine production, correlating with increased Cish expression and decreased TCR signaling. Collectively, our study highlights a critical role for METTL14-dependent-m6A modification in iNKT cell development and function.
Keywords: CD1; CP: Immunology; CP: Molecular biology; NKT cells; T cell development; knockout mice; m(6)A.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests H.H. is an employee of Genentech.
Figures
References
-
- Baranek T, Lebrigand K, de Amat Herbozo C, Gonzalez L, Bogard G, Dietrich C, Magnone V, Boisseau C, Jouan Y, Trottein F, et al. (2020). High dimensional single-cell analysis reveals iNKT cell developmental trajectories and effector fate decision. Cell Rep 32, 108116. 10.1016/j.celrep.2020.108116. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
