

AP-1 transcription factor network explains diverse patterns of cellular plasticity in melanoma cells
- PMID: 35926467
- PMCID: PMC9395172
- DOI: 10.1016/j.celrep.2022.111147
AP-1 transcription factor network explains diverse patterns of cellular plasticity in melanoma cells
Abstract
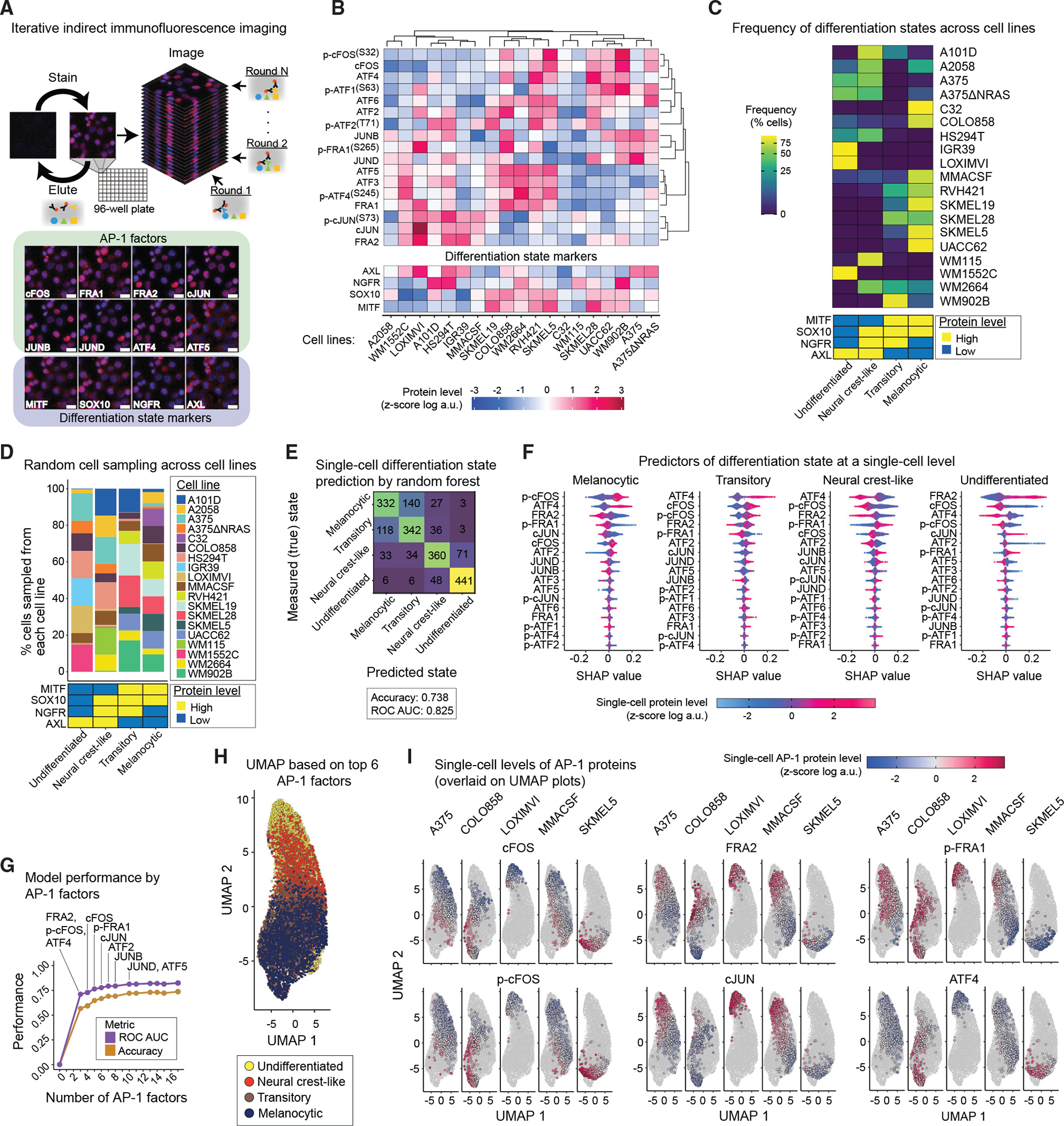
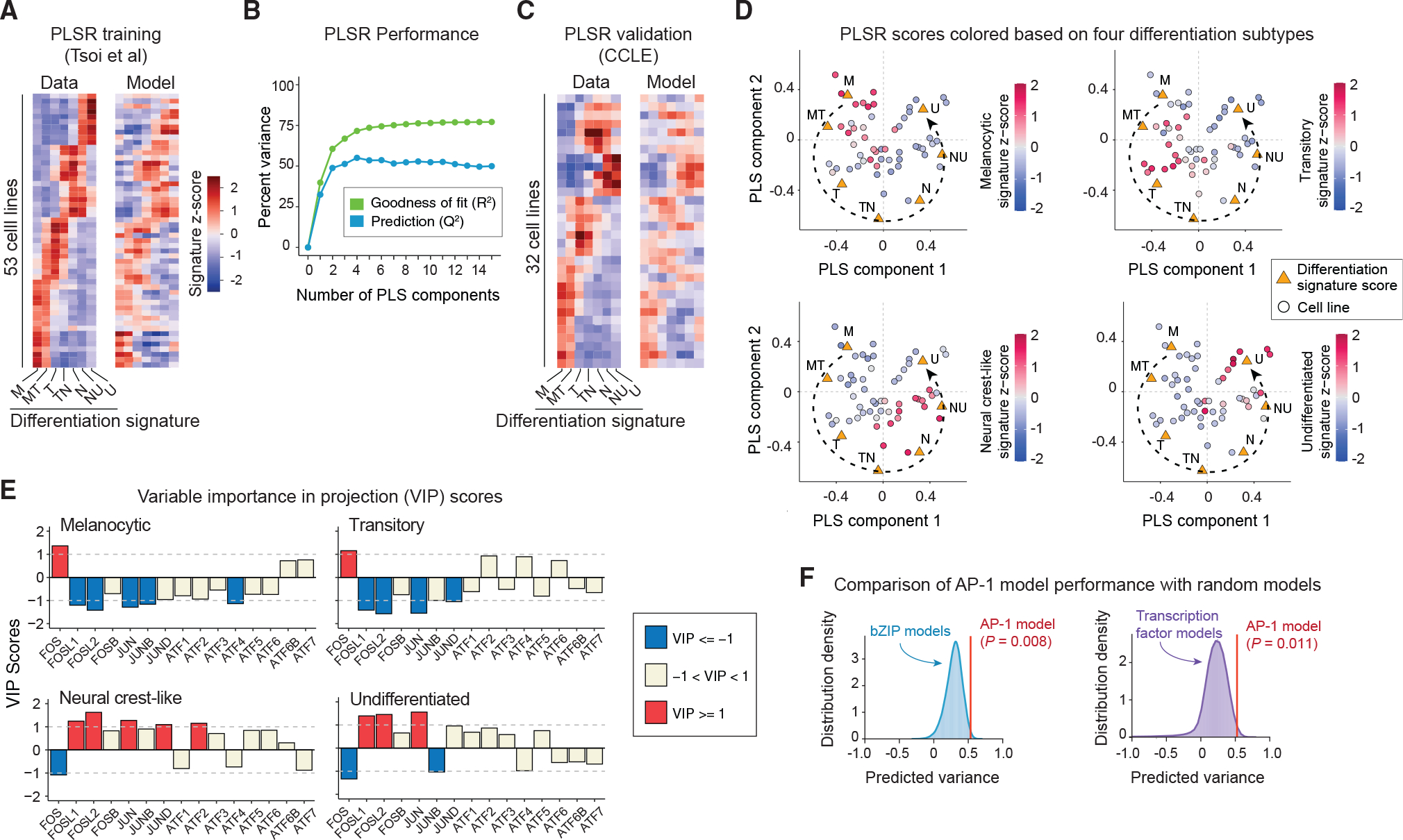
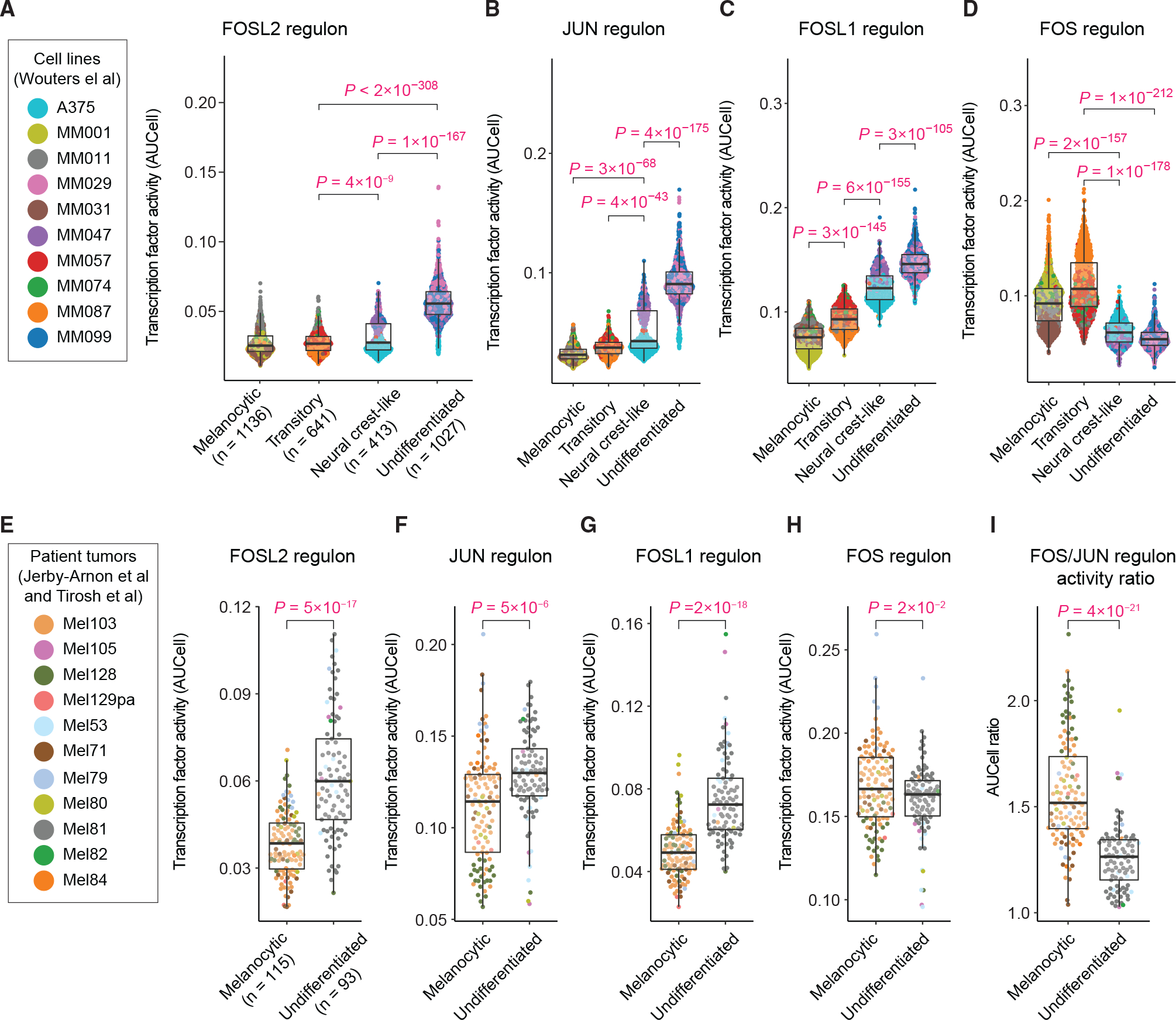
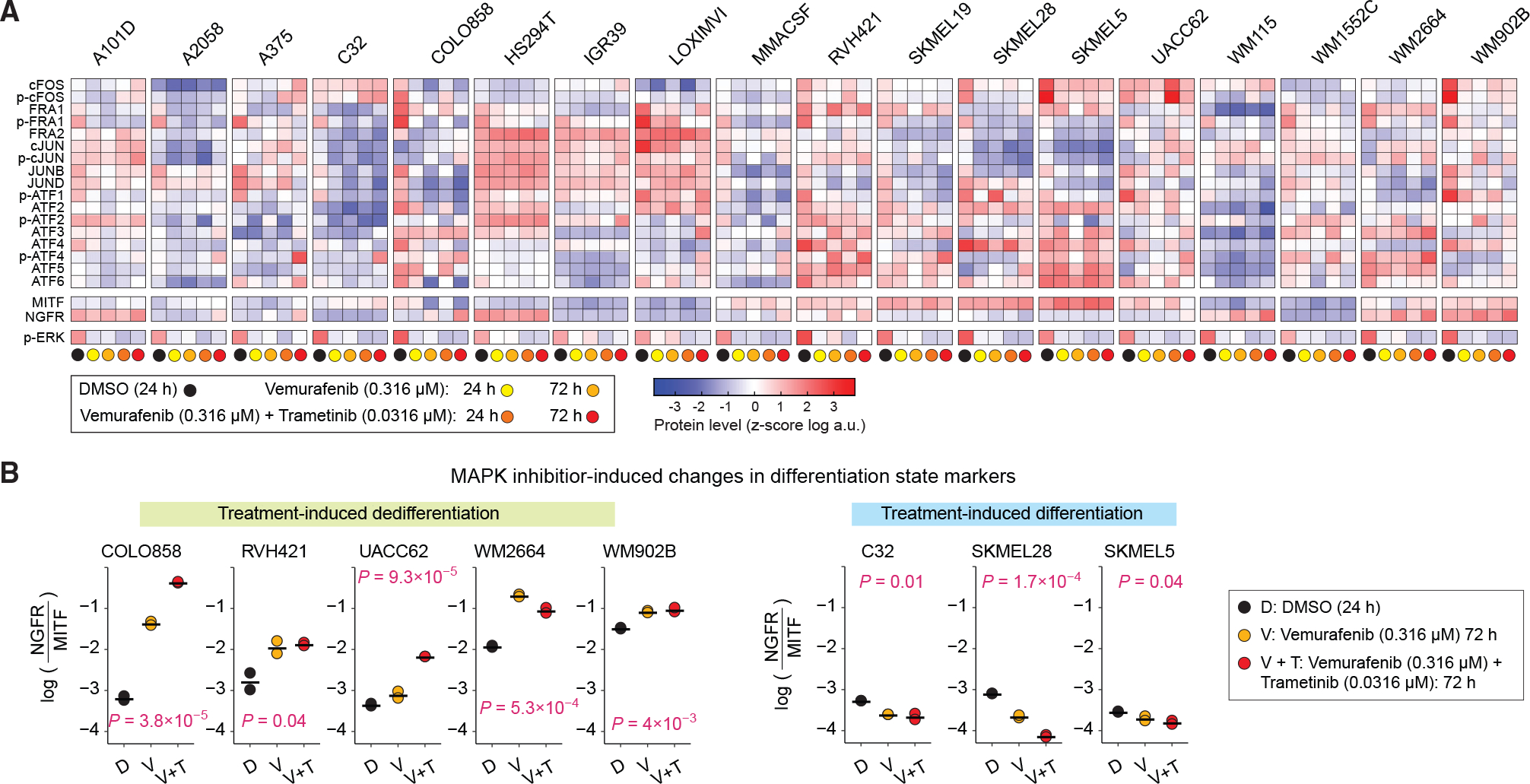
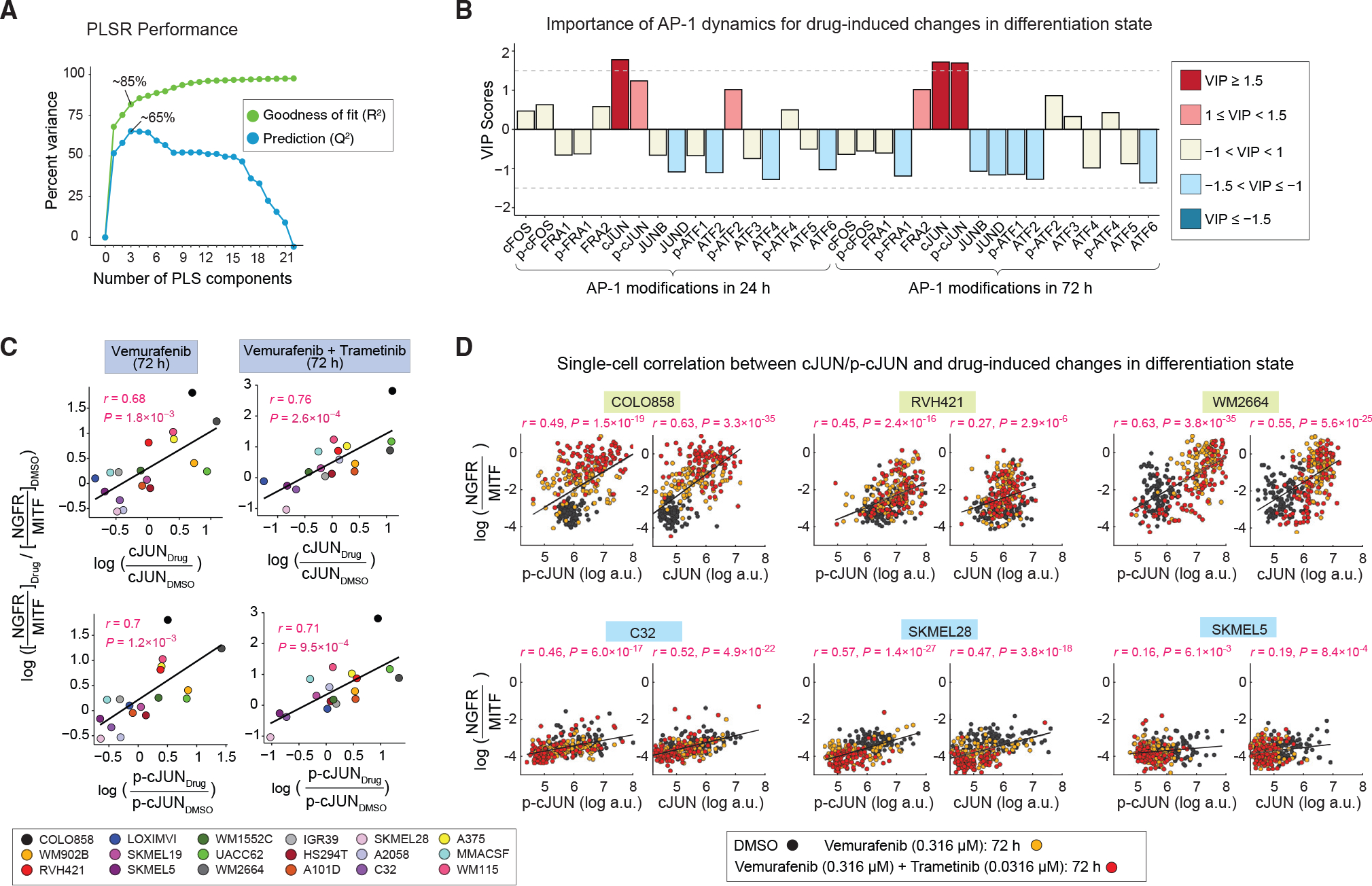
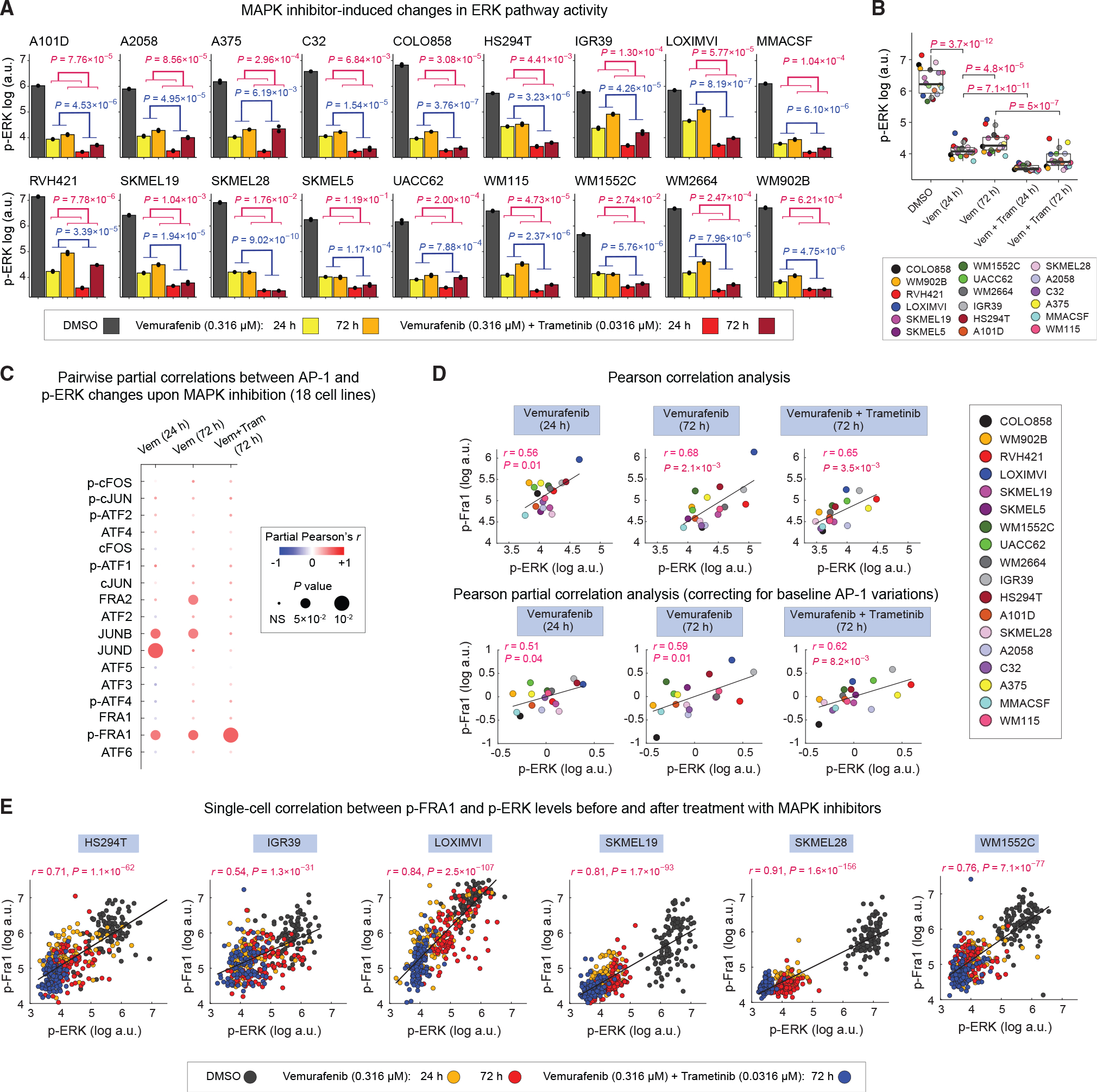
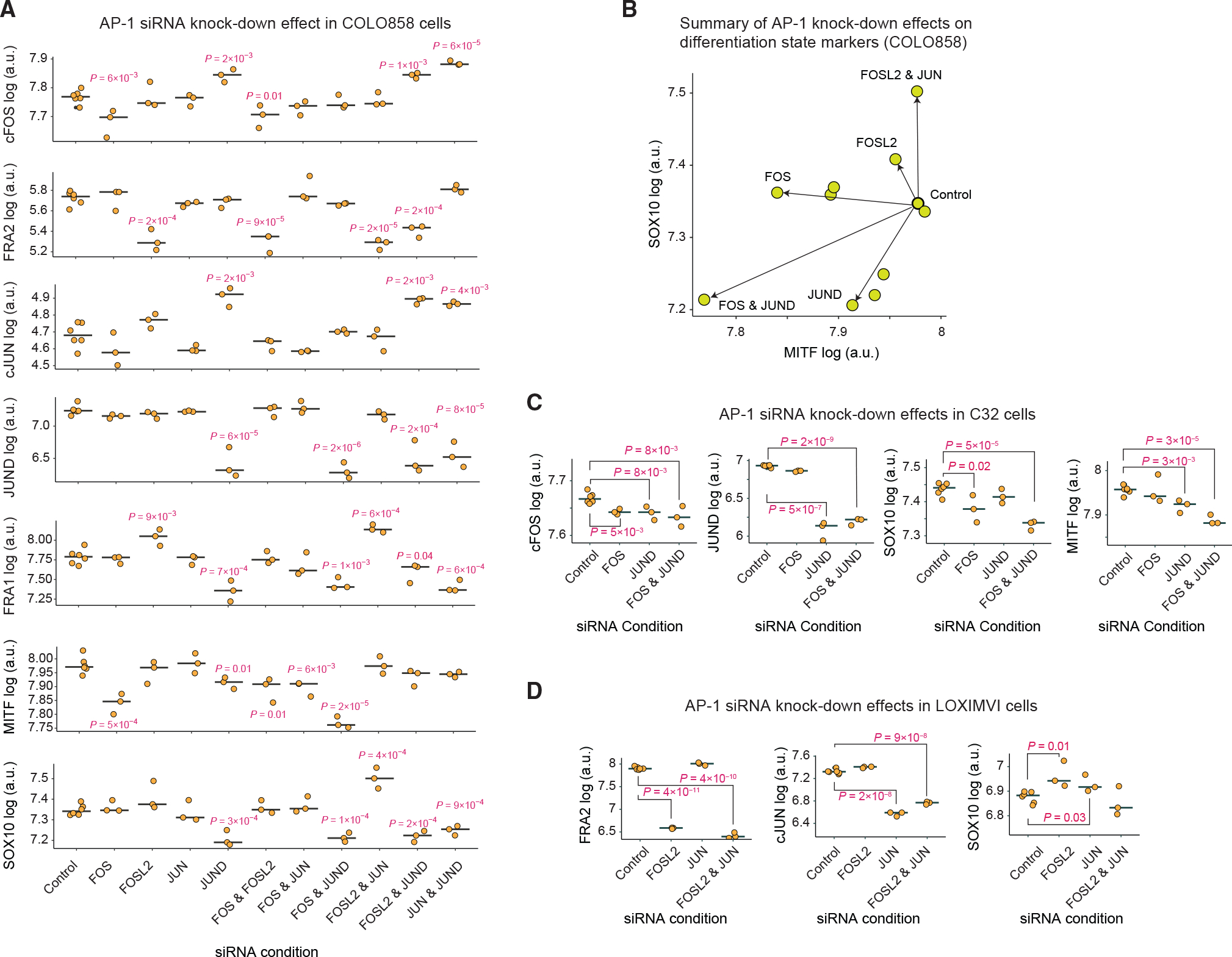
Cellular plasticity associated with fluctuations in transcriptional programs allows individual cells in a tumor to adopt heterogeneous differentiation states and switch phenotype during their adaptive responses to therapies. Despite increasing knowledge of such transcriptional programs, the molecular basis of cellular plasticity remains poorly understood. Here, we combine multiplexed transcriptional and protein measurements at population and single-cell levels with multivariate statistical modeling to show that the state of AP-1 transcription factor network plays a unifying role in explaining diverse patterns of plasticity in melanoma. We find that a regulated balance among AP-1 factors cJUN, JUND, FRA2, FRA1, and cFOS determines the intrinsic diversity of differentiation states and adaptive responses to MAPK inhibitors in melanoma cells. Perturbing this balance through genetic depletion of specific AP-1 proteins, or by MAPK inhibitors, shifts cellular heterogeneity in a predictable fashion. Thus, AP-1 may serve as a critical node for manipulating cellular plasticity with potential therapeutic implications.
Keywords: AP-1 transcription factors; BRAF-mutant melanoma; CP: Cancer; CP: Molecular biology; MAPK signaling pathway; adaptive drug resistance; cellular plasticity; computational modeling; differentiation state heterogeneity; single-cell analysis; statistical learning; targeted therapies.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
