

Neutrophil extracellular traps are involved in enhanced contact hypersensitivity response in IL-36 receptor antagonist-deficient mice
- PMID: 35927298
- PMCID: PMC9352770
- DOI: 10.1038/s41598-022-16449-z
Neutrophil extracellular traps are involved in enhanced contact hypersensitivity response in IL-36 receptor antagonist-deficient mice
Abstract
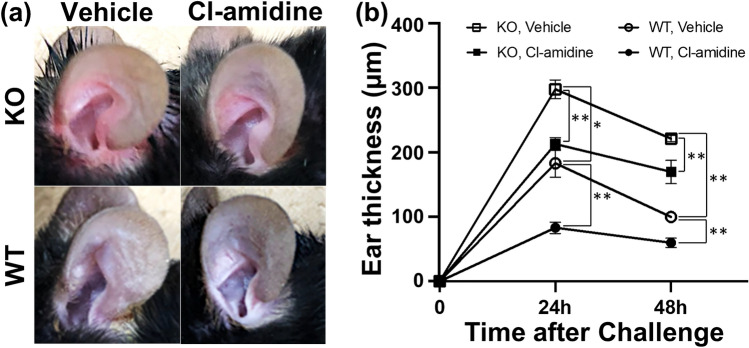
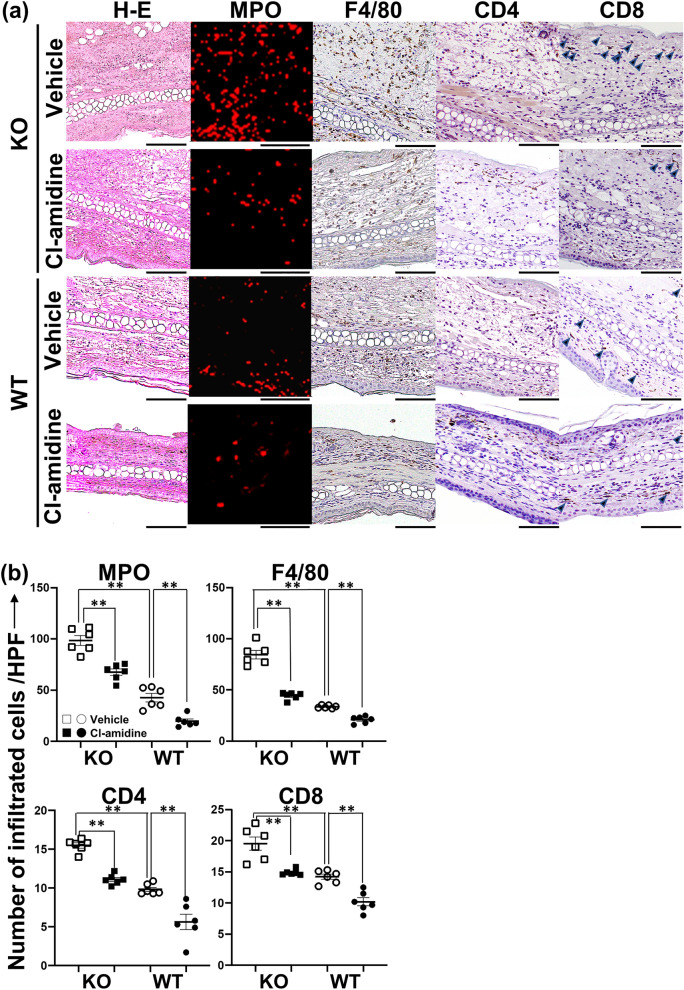
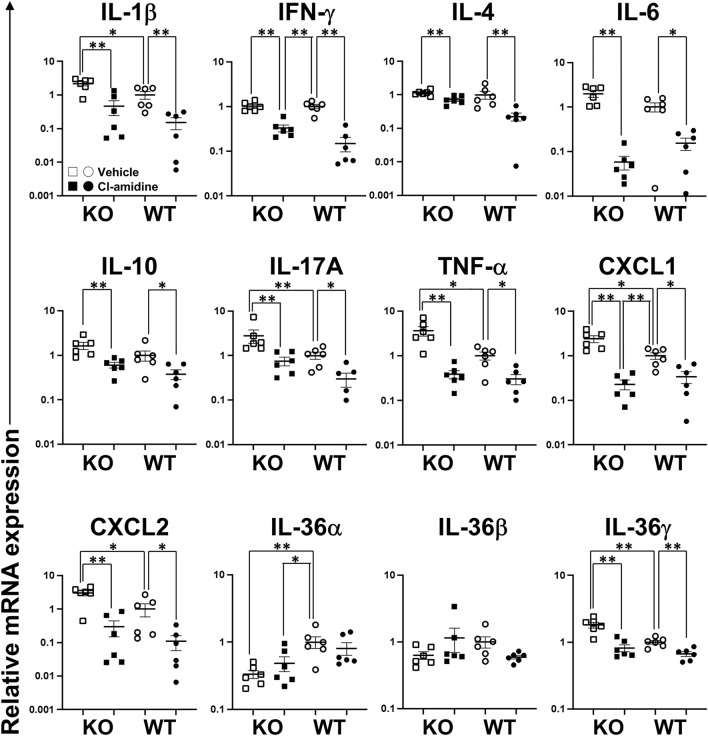
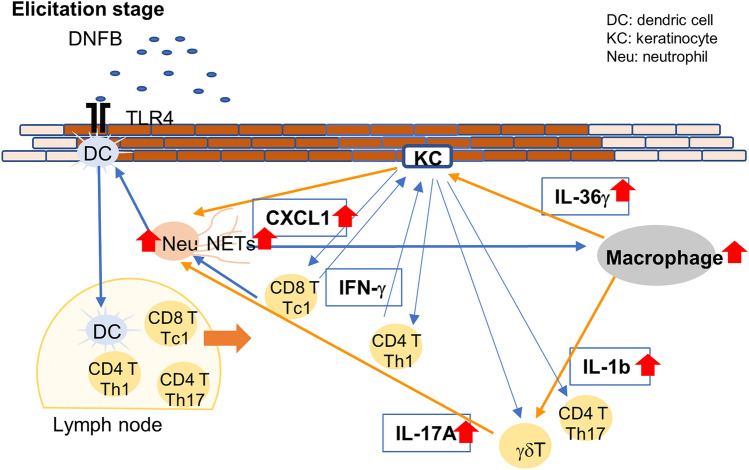
Loss-of-function homozygous or compound heterozygous mutations in IL36RN, which encodes interleukin-36 receptor antagonist (IL-36Ra), have been implicated in the pathogenesis of skin disorders. We previously reported that Il36rn-/- mice exhibit an enhanced contact hypersensitivity (CHS) response through increased neutrophil recruitment. In addition, Il36rn-/- mice show severe imiquimod-induced psoriatic skin lesions and enhanced neutrophil extracellular trap (NET) formation. We hypothesized that NETs may play an important role in the CHS response. To confirm this, we examined the CHS response and NET formation in Il36rn-/- mice. Il36rn-/- mice showed enhanced CHS responses, increased infiltration of inflammatory cells, including neutrophils, CD4+ T cells, and CD8+ T cells, NET formation, and enhanced mRNA expression of cytokines and chemokines, including IL-1β, C-X-C motif chemokine ligand (CXCL)1, CXCL2, and IL-36γ. Furthermore, NET formation blockade improved the CHS response, which consequently decreased inflammatory cell infiltration and NET formation. Consistently, we observed decreased expression of these cytokines and chemokines. These findings indicate that IL-36Ra deficiency aggravates the CHS response caused by excessive inflammatory cell recruitment, NET formation, and cytokine and chemokine production, and that NET formation blockade alleviates the CHS response. Thus, NET formation may play a prominent role in the CHS response.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Cutaneous ischemia-reperfusion injury is exacerbated by IL-36 receptor antagonist deficiency.J Eur Acad Dermatol Venereol. 2022 Feb;36(2):295-304. doi: 10.1111/jdv.17767. Epub 2021 Nov 20. J Eur Acad Dermatol Venereol. 2022. PMID: 34699104
-
TAK-242 ameliorates contact dermatitis exacerbated by IL-36 receptor antagonist deficiency.Sci Rep. 2020 Jan 20;10(1):734. doi: 10.1038/s41598-020-57550-5. Sci Rep. 2020. PMID: 31959814 Free PMC article.
-
Neutrophil extracellular traps are induced in a psoriasis model of interleukin-36 receptor antagonist-deficient mice.Sci Rep. 2020 Nov 19;10(1):20149. doi: 10.1038/s41598-020-76864-y. Sci Rep. 2020. PMID: 33214582 Free PMC article.
-
IL-36 receptor antagonist deficiency resulted in delayed wound healing due to excessive recruitment of immune cells.Sci Rep. 2020 Sep 8;10(1):14772. doi: 10.1038/s41598-020-71256-8. Sci Rep. 2020. PMID: 32901055 Free PMC article.
-
Neutrophil expression of Fas ligand and perforin directs effector CD8 T cell infiltration into antigen-challenged skin.J Immunol. 2012 Sep 1;189(5):2191-202. doi: 10.4049/jimmunol.1102729. Epub 2012 Jul 18. J Immunol. 2012. PMID: 22815291 Free PMC article.
Cited by
-
Plasma Levels of Interleukins 36α, 36β, and 37 in Patients with Psoriasis and Their Correlation with Disease Activity Parameters.J Clin Med. 2022 Sep 6;11(18):5254. doi: 10.3390/jcm11185254. J Clin Med. 2022. PMID: 36142901 Free PMC article.
-
Role of Innate Immunity in Allergic Contact Dermatitis: An Update.Int J Mol Sci. 2023 Aug 19;24(16):12975. doi: 10.3390/ijms241612975. Int J Mol Sci. 2023. PMID: 37629154 Free PMC article. Review.
-
Role of IL-1 Family Cytokines IL-36, IL-37, IL-38 in Osteoarthritis and Rheumatoid Arthritis: A Comprehensive Review.J Inflamm Res. 2024 Jun 20;17:4001-4016. doi: 10.2147/JIR.S474879. eCollection 2024. J Inflamm Res. 2024. PMID: 38915806 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
