

Global phosphoproteomic analysis identified key kinases regulating male meiosis in mouse
- PMID: 35930080
- PMCID: PMC11071816
- DOI: 10.1007/s00018-022-04507-8
Global phosphoproteomic analysis identified key kinases regulating male meiosis in mouse
Abstract
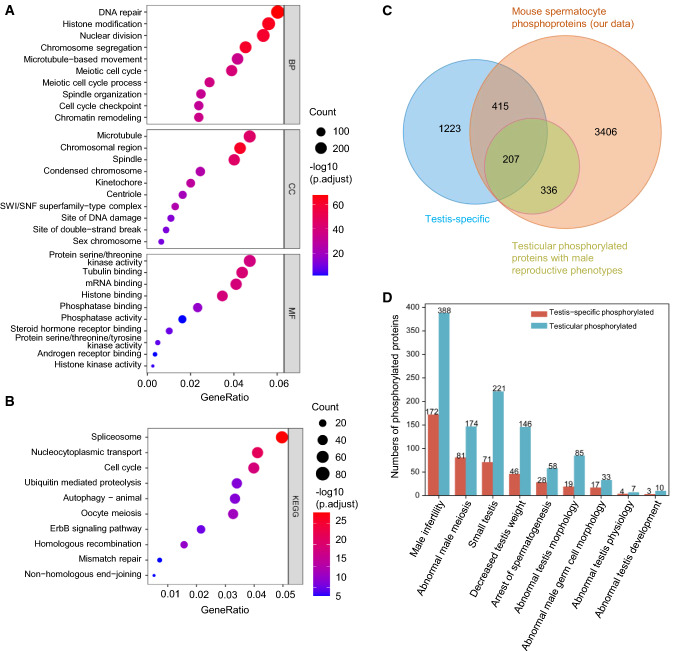
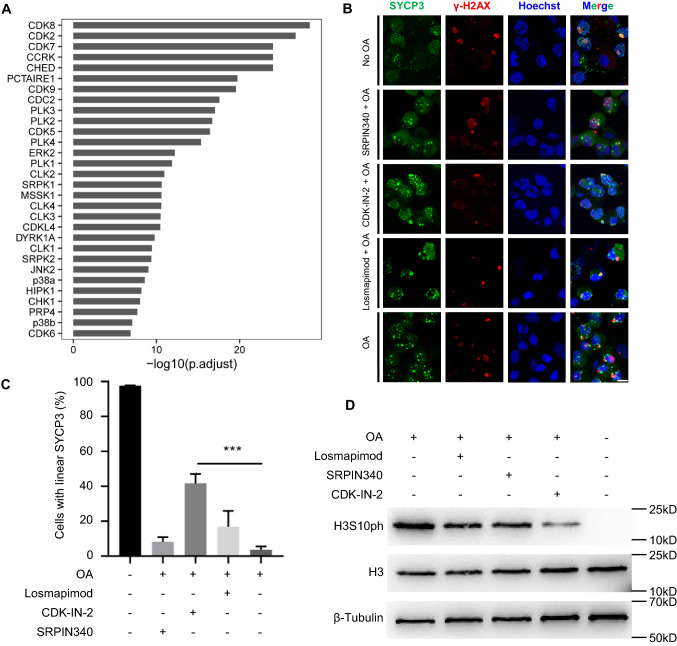
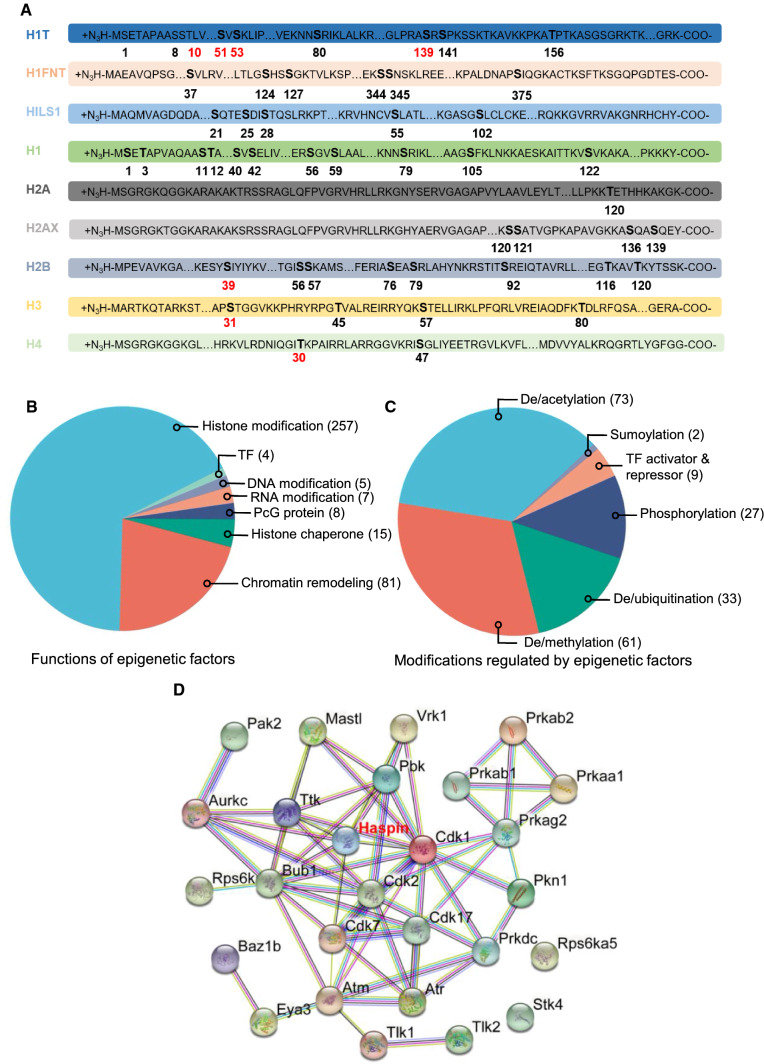
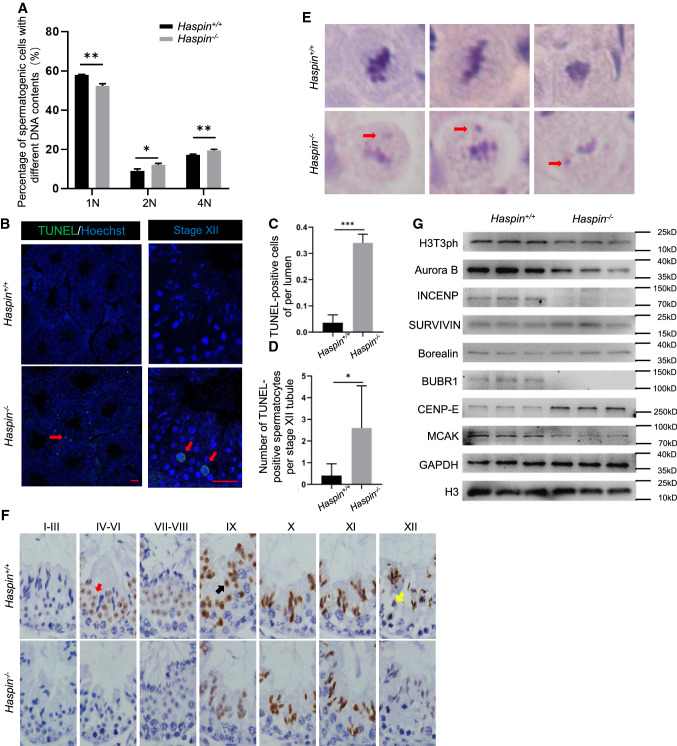
Meiosis, a highly conserved process in organisms from fungi to mammals, is subjected to protein phosphorylation regulation. Due to the low abundance of phosphorylation, there is a lack of systemic characterization of phosphorylation regulation of meiosis in mammals. Using the phosphoproteomic approach, we profiled large-scale phosphoproteome of purified primary spermatocytes undergoing meiosis I, and identified 14,660 phosphorylation sites in 4419 phosphoproteins. Kinase-substrate phosphorylation network analysis followed by in vitro meiosis study showed that CDK9 was essential for meiosis progression to metaphase I and had enriched substrate phosphorylation sites in proteins involved in meiotic cell cycle. In addition, histones and epigenetic factors were found to be widely phosphorylated. Among those, HASPIN was found to be essential for male fertility. Haspin knockout led to misalignment of chromosomes, apoptosis of metaphase spermatocytes and a decreased number of sperm by deregulation of H3T3ph, chromosomal passenger complex (CPC) and spindle assembly checkpoint (SAC). The complicated protein phosphorylation and its important regulatory functions in meiosis indicated that in-depth studies of phosphorylation-mediated signaling could help us elucidate the mechanisms of meiosis.
Keywords: CDK9; Chromosomal passenger complex; Haspin; Histone phosphorylation; Phosphoproteome; Spermatocyte; Spindle assembly checkpoint.
© 2022. The Author(s), under exclusive licence to Springer Nature Switzerland AG.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
