

Proteogenomics reveals sex-biased aging genes and coordinated splicing in cardiac aging
- PMID: 35930447
- PMCID: PMC9448281
- DOI: 10.1152/ajpheart.00244.2022
Proteogenomics reveals sex-biased aging genes and coordinated splicing in cardiac aging
Abstract
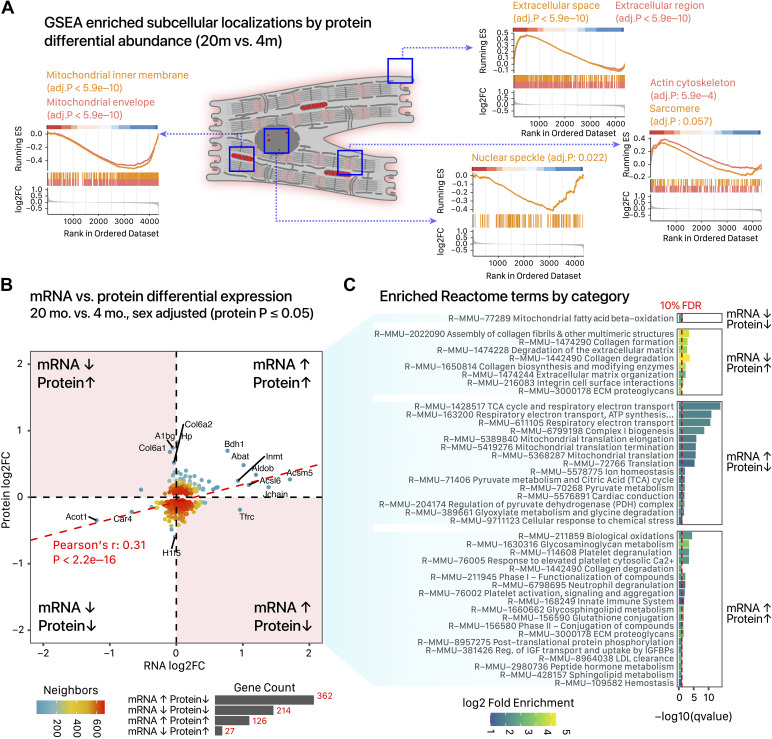
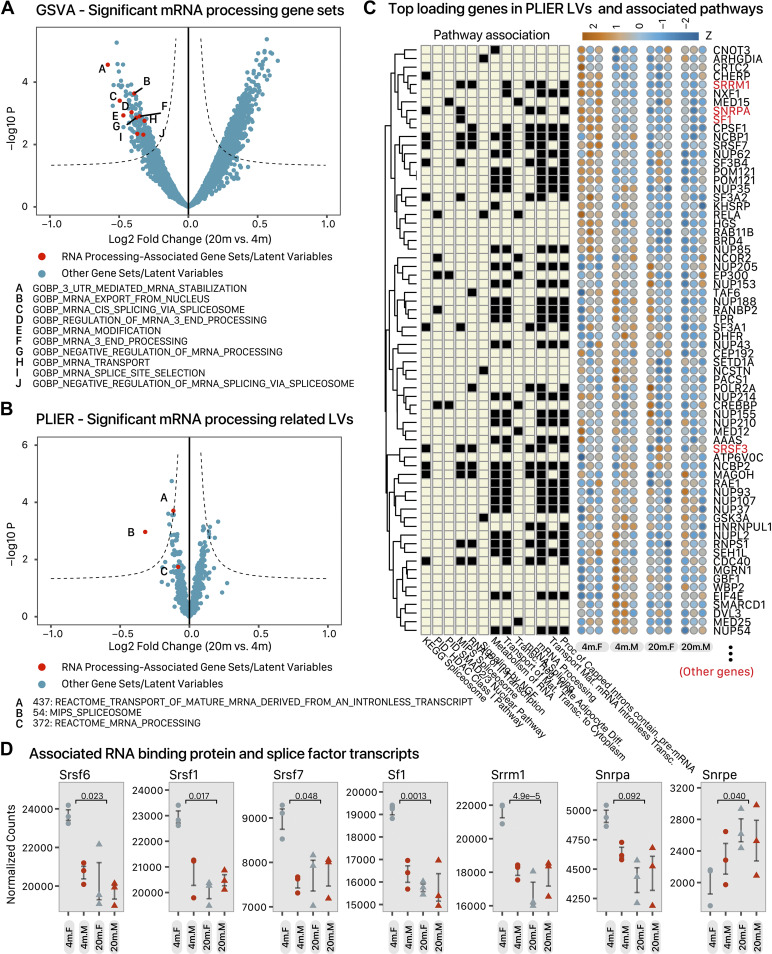
The risks of heart diseases are significantly modulated by age and sex, but how these factors influence baseline cardiac gene expression remains incompletely understood. Here, we used RNA sequencing and mass spectrometry to compare gene expression in female and male young adult (4 mo) and early aging (20 mo) mouse hearts, identifying thousands of age- and sex-dependent gene expression signatures. Sexually dimorphic cardiac genes are broadly distributed, functioning in mitochondrial metabolism, translation, and other processes. In parallel, we found over 800 genes with differential aging response between male and female, including genes in cAMP and PKA signaling. Analysis of the sex-adjusted aging cardiac transcriptome revealed a widespread remodeling of exon usage patterns that is largely independent from differential gene expression, concomitant with upstream changes in RNA-binding protein and splice factor transcripts. To evaluate the impact of the splicing events on cardiac proteoform composition, we applied an RNA-guided proteomics computational pipeline to analyze the mass spectrometry data and detected hundreds of putative splice variant proteins that have the potential to rewire the cardiac proteome. Taken together, the results here suggest that cardiac aging is associated with 1) widespread sex-biased aging genes and 2) a rewiring of RNA splicing programs, including sex- and age-dependent changes in exon usages and splice patterns that have the potential to influence cardiac protein structure and function. These changes contribute to the emerging evidence for considerable sexual dimorphism in the cardiac aging process that should be considered in the search for disease mechanisms.NEW & NOTEWORTHY Han et al. used proteogenomics to compare male and female mouse hearts at 4 and 20 mo. Sex-biased cardiac genes function in mitochondrial metabolism, translation, autophagy, and other processes. Hundreds of cardiac genes show sex-by-age interactions, that is, sex-biased aging genes. Cardiac aging is accompanied with a remodeling of exon usage in functionally coordinated genes, concomitant with differential expression of RNA-binding proteins and splice factors. These features represent an underinvestigated aspect of cardiac aging that may be relevant to the search for disease mechanisms.
Keywords: aging; alternative splicing; proteoforms; proteogenomics; sex difference.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures





Comment in
-
The big picture: cardiac sex-age interactions and proteogenomic insights.Am J Physiol Heart Circ Physiol. 2022 Oct 1;323(4):H640-H642. doi: 10.1152/ajpheart.00418.2022. Epub 2022 Sep 9. Am J Physiol Heart Circ Physiol. 2022. PMID: 36083794 No abstract available.
References
-
- Cheng S, Xanthakis V, Sullivan LM, Lieb W, Massaro J, Aragam J, Benjamin EJ, Vasan RS. Correlates of echocardiographic indices of cardiac remodeling over the adult life course: longitudinal observations from the Framingham Heart Study. Circulation 122: 570–578, 2010. doi: 10.1161/CIRCULATIONAHA.110.937821. - DOI - PMC - PubMed
-
- Sotomi Y, Hikoso S, Nakatani D, Mizuno H, Okada K, Dohi T, Kitamura T, Sunaga A, Kida H, Oeun B, Sato T, Komukai S, Tamaki S, Yano M, Hayashi T, Nakagawa A, Nakagawa Y, Yasumura Y, Yamada T, Sakata Y; PURSUIT‐HFpEF Investigators. Sex differences in heart failure with preserved ejection fraction. J Am Heart Assoc 10: e018574, 2021. doi: 10.1161/JAHA.120.018574. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
- R21-HL150456/HHS | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R00-HL144829/HHS | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R00 HL127302/HL/NHLBI NIH HHS/United States
- R03-OD032666/HHS | NIH | NIH Office of the Director (OD)
- R01 HL141278/HL/NHLBI NIH HHS/United States
- F32 HL149191/HL/NHLBI NIH HHS/United States
- F32-HL149191/HHS | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R00-HL127302/HHS | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R21 HL150456/HL/NHLBI NIH HHS/United States
- R03 OD032666/OD/NIH HHS/United States
- R00 HL144829/HL/NHLBI NIH HHS/United States
- R01-HL141278/HHS | NIH | National Heart, Lung, and Blood Institute (NHLBI)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases