

lncRNA-GM targets Foxo1 to promote T cell-mediated autoimmunity
- PMID: 35930633
- PMCID: PMC9355365
- DOI: 10.1126/sciadv.abn9181
lncRNA-GM targets Foxo1 to promote T cell-mediated autoimmunity
Abstract
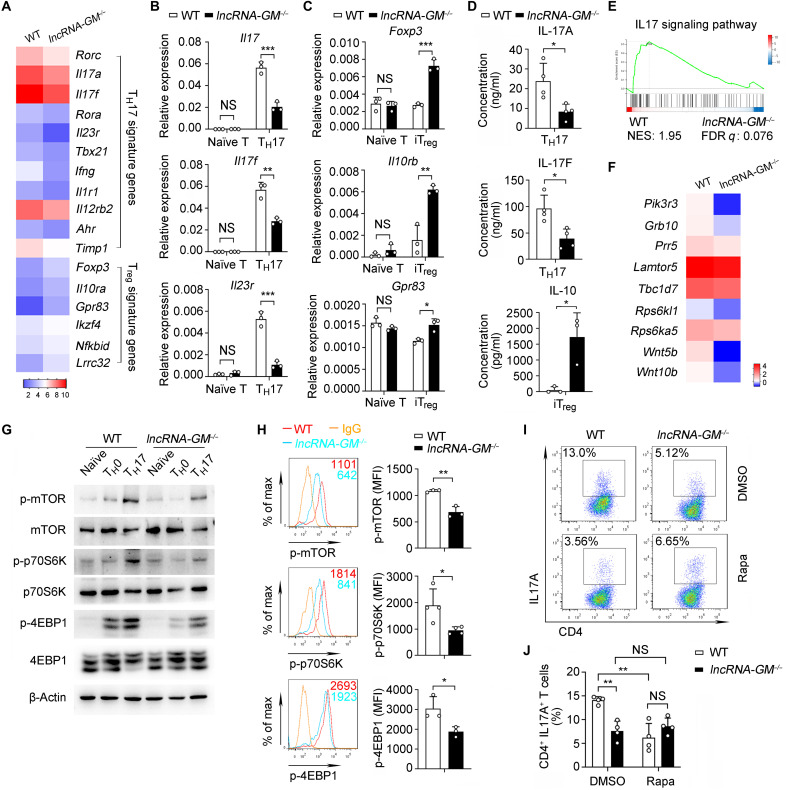
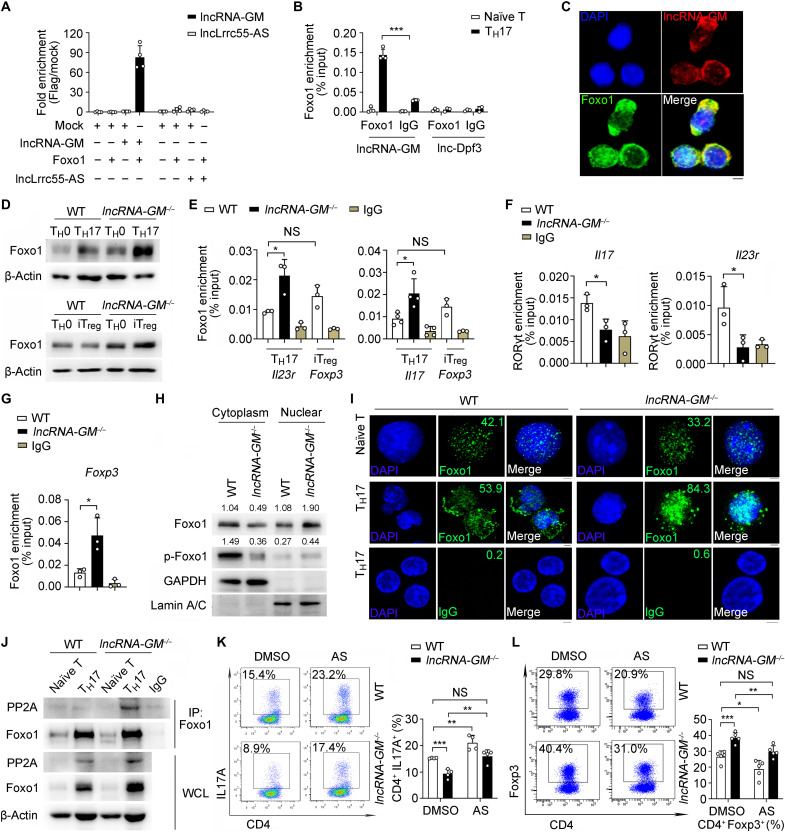
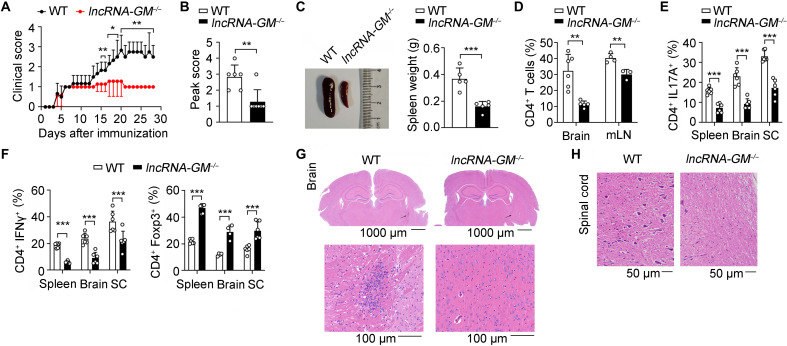
RNA-RBP interaction is important in immune regulation and implicated in various immune disorders. The differentiation of proinflammatory T cell subset TH17 and its balance with regulatory T cell (Treg) generation is closely related to autoimmune pathogenesis. The roles of RNA-RBP interaction in regulation of TH17/Treg differentiation and autoinflammation remain in need of further investigation. Here we report that lncRNA-GM polarizes TH17 differentiation but inhibits iTreg differentiation by reducing activity of Foxo1, a transcriptional factor that is important in inhibiting TH17 differentiation but promoting Treg generation. lncRNA-GM-deficient mice were protected from experimental autoimmune encephalomyelitis. Mechanistically, lncRNA-GM directly binds to cytoplasmic Foxo1, thus inhibiting its activity through blocking dephosphorylation of Foxo1 by phosphatase PP2A to promote Il23r transcription. The human homolog of lncRNA-GM (AK026392.1) also polarizes human TH17 differentiation. Our study provides mechanistic insight into the interaction of lncRNA and transcriptional factor in determining T cell subset differentiation during T cell-mediated autoimmune diseases.
Figures



References
-
- Fugger L., Jensen L. T., Rossjohn J., Challenges, progress, and prospects of developing therapies to treat autoimmune diseases. Cell 181, 63–80 (2020). - PubMed
-
- Chataway J., Tackling progression in multiple sclerosis. Lancet Neurol. 17, 489–491 (2018). - PubMed
-
- Ghoreschi K., Laurence A., Yang X.-P., Tato C. M., McGeachy M. J., Konkel J. E., Ramos H. L., Wei L., Davidson T. S., Bouladoux N., Grainger J. R., Chen Q., Kanno Y., Watford W. T., Sun H.-W., Eberl G., Shevach E. M., Belkaid Y., Cua D. J., Chen W., O’Shea J. J., Generation of pathogenic TH17 cells in the absence of TGF-β signalling. Nature 467, 967–971 (2010). - PMC - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
