

The duper mutation reveals previously unsuspected functions of Cryptochrome 1 in circadian entrainment and heart disease
- PMID: 35930669
- PMCID: PMC9371649
- DOI: 10.1073/pnas.2121883119
The duper mutation reveals previously unsuspected functions of Cryptochrome 1 in circadian entrainment and heart disease
Abstract
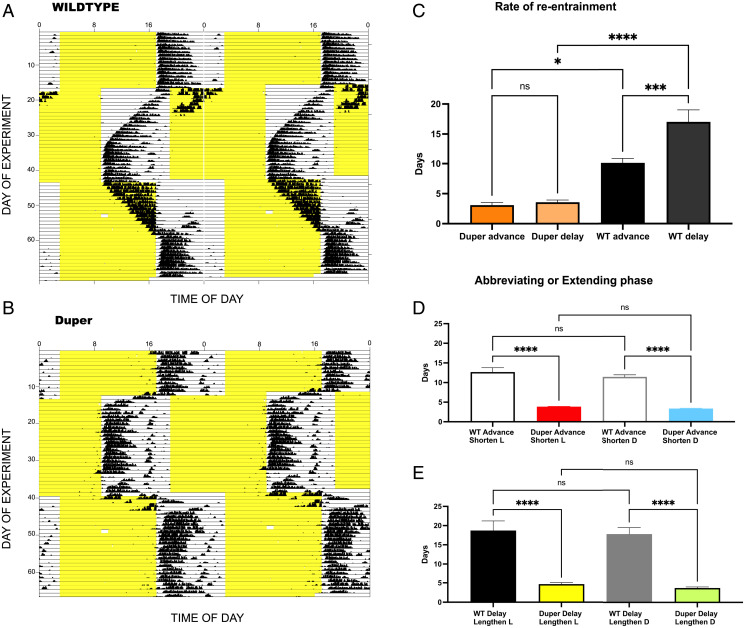
The Cryptochrome 1 (Cry1)-deficient duper mutant hamster has a short free-running period in constant darkness (τDD) and shows large phase shifts in response to brief light pulses. We tested whether this measure of the lability of the circadian phase is a general characteristic of Cry1-null animals and whether it indicates resistance to jet lag. Upon advance of the light:dark (LD) cycle, both duper hamsters and Cry1-/- mice re-entrained locomotor rhythms three times as fast as wild types. However, accelerated re-entrainment was dissociated from the amplified phase-response curve (PRC): unlike duper hamsters, Cry1-/- mice show no amplification of the phase response to 15' light pulses. Neither the amplified acute shifts nor the increased rate of re-entrainment in duper mutants is due to acceleration of the circadian clock: when mutants drank heavy water to lengthen the period, these aspects of the phenotype persisted. In light of the health consequences of circadian misalignment, we examined effects of duper and phase shifts on a hamster model of heart disease previously shown to be aggravated by repeated phase shifts. The mutation shortened the lifespan of cardiomyopathic hamsters relative to wild types, but this effect was eliminated when mutants experienced 8-h phase shifts every second week, to which they rapidly re-entrained. Our results reveal previously unsuspected roles of Cry1 in phase shifting and longevity in the face of heart disease. The duper mutant offers new opportunities to understand the basis of circadian disruption and jet lag.
Keywords: Cryptochrome 1; cardiomyopathy; circadian rhythms; entrainment; jet lag.
Conflict of interest statement
The authors declare no competing interest.
Figures



Similar articles
-
Adult Neurogenesis Is Altered by Circadian Phase Shifts and the Duper Mutation in Female Syrian Hamsters.eNeuro. 2023 Mar 29;10(3):ENEURO.0359-22.2023. doi: 10.1523/ENEURO.0359-22.2023. Print 2023 Mar. eNeuro. 2023. PMID: 36878716 Free PMC article.
-
Effects of the duper mutation on responses to light: parametric and nonparametric responses, range of entrainment, and masking.J Biol Rhythms. 2014 Apr;29(2):97-109. doi: 10.1177/0748730413520399. J Biol Rhythms. 2014. PMID: 24682204
-
Phase resetting in duper hamsters: specificity to photic zeitgebers and circadian phase.J Biol Rhythms. 2015 Apr;30(2):129-43. doi: 10.1177/0748730414568297. Epub 2015 Jan 29. J Biol Rhythms. 2015. PMID: 25633984 Free PMC article.
-
From circadian clock mechanism to sleep disorders and jet lag: Insights from a computational approach.Biochem Pharmacol. 2021 Sep;191:114482. doi: 10.1016/j.bcp.2021.114482. Epub 2021 Feb 20. Biochem Pharmacol. 2021. PMID: 33617843 Review.
-
Modeling the circadian clock: from molecular mechanism to physiological disorders.Bioessays. 2008 Jun;30(6):590-600. doi: 10.1002/bies.20762. Bioessays. 2008. PMID: 18478538 Review.
Cited by
-
Adult Neurogenesis Is Altered by Circadian Phase Shifts and the Duper Mutation in Female Syrian Hamsters.eNeuro. 2023 Mar 29;10(3):ENEURO.0359-22.2023. doi: 10.1523/ENEURO.0359-22.2023. Print 2023 Mar. eNeuro. 2023. PMID: 36878716 Free PMC article.
-
Neuroendocrine effects of the duper mutation in Syrian hamsters: a role for Cryptochrome 1.Front Physiol. 2024 Feb 20;15:1351682. doi: 10.3389/fphys.2024.1351682. eCollection 2024. Front Physiol. 2024. PMID: 38444761 Free PMC article.
-
duper is a null mutation of Cryptochrome 1 in Syrian hamsters.Proc Natl Acad Sci U S A. 2022 May 3;119(18):e2123560119. doi: 10.1073/pnas.2123560119. Epub 2022 Apr 26. Proc Natl Acad Sci U S A. 2022. PMID: 35471909 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
