

A new fluorescent sensor mitoferrofluor indicates the presence of chelatable iron in polarized and depolarized mitochondria
- PMID: 35931111
- PMCID: PMC9460511
- DOI: 10.1016/j.jbc.2022.102336
A new fluorescent sensor mitoferrofluor indicates the presence of chelatable iron in polarized and depolarized mitochondria
Abstract
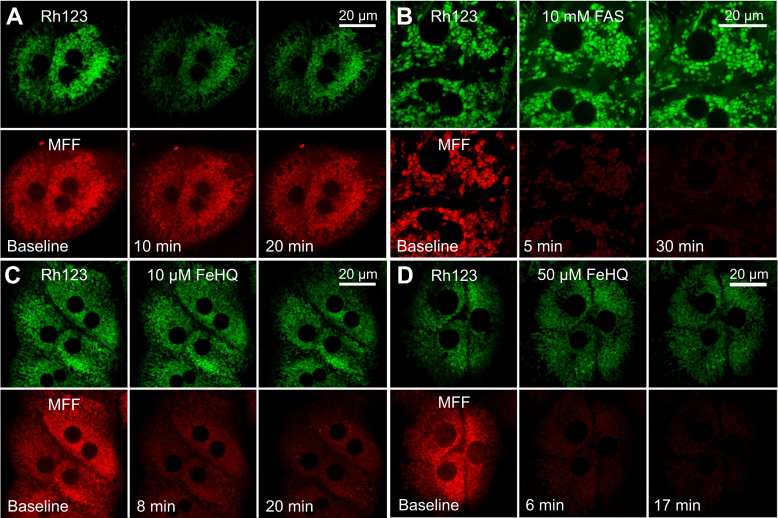
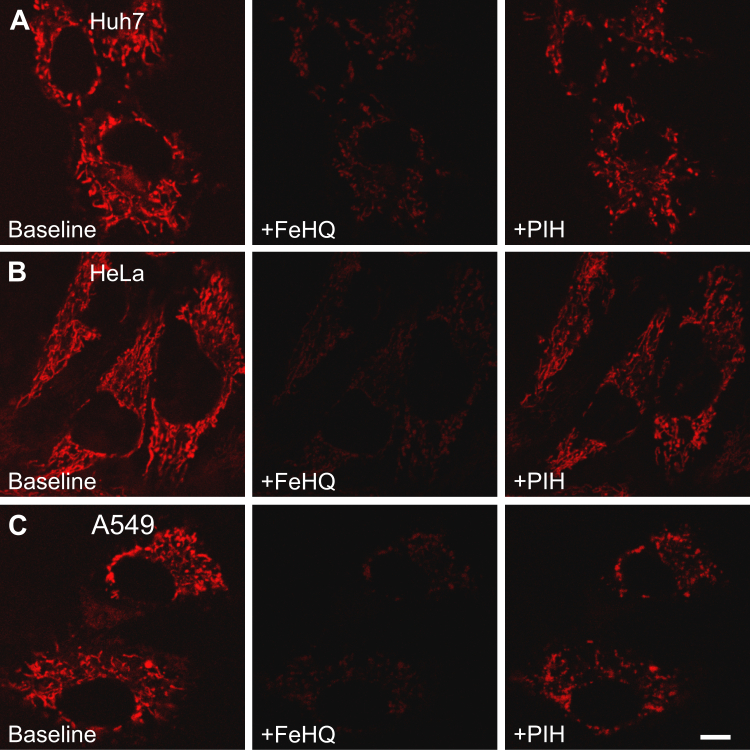
Mitochondrial chelatable iron contributes to the severity of several injury processes, including ischemia/reperfusion, oxidative stress, and drug toxicity. However, methods to measure this species in living cells are lacking. To measure mitochondrial chelatable iron in living cells, here we synthesized a new fluorescent indicator, mitoferrofluor (MFF). We designed cationic MFF to accumulate electrophoretically in polarized mitochondria, where a reactive group then forms covalent adducts with mitochondrial proteins to retain MFF even after subsequent depolarization. We also show in cell-free medium that Fe2+ (and Cu2+), but not Fe3+, Ca2+, or other biologically relevant divalent cations, strongly quenched MFF fluorescence. Using confocal microscopy, we demonstrate in hepatocytes that red MFF fluorescence colocalized with the green fluorescence of the mitochondrial membrane potential (ΔΨm) indicator, rhodamine 123 (Rh123), indicating selective accumulation into the mitochondria. Unlike Rh123, mitochondria retained MFF after ΔΨm collapse. Furthermore, intracellular delivery of iron with membrane-permeant Fe3+/8-hydroxyquinoline (FeHQ) quenched MFF fluorescence by ∼80% in hepatocytes and other cell lines, which was substantially restored by the membrane-permeant transition metal chelator pyridoxal isonicotinoyl hydrazone. We also show FeHQ quenched the fluorescence of cytosolically coloaded calcein, another Fe2+ indicator, confirming that Fe3+ in FeHQ undergoes intracellular reduction to Fe2+. Finally, MFF fluorescence did not change after addition of the calcium mobilizer thapsigargin, which shows MFF is insensitive to physiologically relevant increases of mitochondrial Ca2+. In conclusion, the new sensor reagent MFF fluorescence is an indicator of mitochondrial chelatable Fe2+ in normal hepatocytes with polarized mitochondria as well as in cells undergoing loss of ΔΨm.
Keywords: iron sensor; ischemia/reperfusion; membrane potential; mitochondria.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures






References
-
- Starke P.E., Farber J.L. Ferric iron and superoxide ions are required for the killing of cultured hepatocytes by hydrogen peroxide. Evidence for the participation of hydroxyl radicals formed by an iron-catalyzed Haber-Weiss reaction. J. Biol. Chem. 1985;260:10099–10104. - PubMed
-
- Kehrer J.P. The Haber-Weiss reaction and mechanisms of toxicity. Toxicology. 2000;149:43–50. - PubMed
-
- Lemasters J.J. In: The Liver. Biology and Pathobiology. Arias I.M., Boyer J.L., Chisari F.V., Fausto N., Schachter D., Shafritz D.A., editors. Lippincott, Williams & Wilkins; Philadelphia, PA: 2001. Hypoxic, ischemic and reperfusion injury to liver; pp. 257–279.
Publication types
MeSH terms
Substances
Grants and funding
- S10 OD018113/OD/NIH HHS/United States
- P30 DK123704/DK/NIDDK NIH HHS/United States
- P20 GM109091/GM/NIGMS NIH HHS/United States
- P30 DK034987/DK/NIDDK NIH HHS/United States
- R01 AA021191/AA/NIAAA NIH HHS/United States
- P30 CA138313/CA/NCI NIH HHS/United States
- T32 DK083262/DK/NIDDK NIH HHS/United States
- P20 GM130457/GM/NIGMS NIH HHS/United States
- R01 DK119523/DK/NIDDK NIH HHS/United States
- R01 AA025379/AA/NIAAA NIH HHS/United States
- P30 GM140964/GM/NIGMS NIH HHS/United States
- UL1 TR001450/TR/NCATS NIH HHS/United States
- R21 ES031335/ES/NIEHS NIH HHS/United States
- R01 DK073336/DK/NIDDK NIH HHS/United States
- R21 AA022815/AA/NIAAA NIH HHS/United States
- P30 DK056350/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Miscellaneous
