

Deficiency of Acetyl-CoA Carboxylase Impairs Digestion, Lipid Synthesis, and Reproduction in the Kissing Bug Rhodnius prolixus
- PMID: 35936892
- PMCID: PMC9353303
- DOI: 10.3389/fphys.2022.934667
Deficiency of Acetyl-CoA Carboxylase Impairs Digestion, Lipid Synthesis, and Reproduction in the Kissing Bug Rhodnius prolixus
Abstract
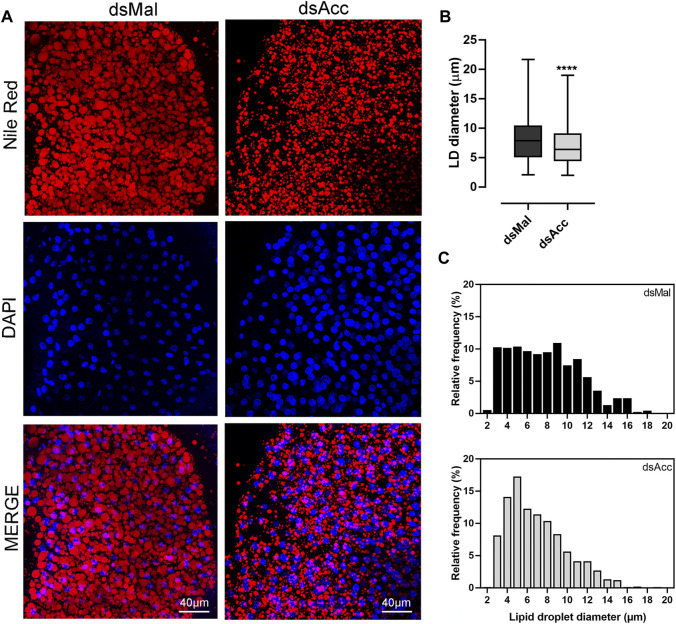
Rhodnius prolixus is a hematophagous insect, vector of Chagas disease. After feeding, as blood is slowly digested, amino acids are used as substrates to fuel lipid synthesis, and adult females accumulate lipids in the fat body and produce eggs. In order to evaluate the importance of de novo fatty acid synthesis for this insect metabolism, we generated acetyl-CoA carboxylase (ACC) deficient insects. The knockdown (AccKD) females had delayed blood digestion and a shorter lifespan. Their fat bodies showed reduced de novo lipogenesis activity, did not accumulate triacylglycerol during the days after blood meal, and had smaller lipid droplets. At 10 days after feeding, there was a general decrease in the amounts of neutral lipids and phospholipids in the fat body. In the hemolymph, no difference was observed in lipid composition at 5 days after blood meal, but at day ten, there was an increase in hydrocarbon content and a decrease in phospholipids. Total protein concentration and amino acid composition were not affected. The AccKD females laid 60% fewer eggs than the control ones, and only 7% hatched (89% for control), although their total protein and triacylglycerol contents were not different. Scanning electron microscopy of the egg surface showed that chorion (eggshell) from the eggs laid by the AccKD insects had an altered ultrastructural pattern when compared to control ones. These results show that ACC has a central role in R. prolixus nutrient homeostasis, and its appropriate activity is important to digestion, lipid synthesis and storage, and reproductive success.
Keywords: Rhodnius prolixus; acetyl-CoA carboxylase (ACC); de novo lipogenesis; fat body; oogenesis.
Copyright © 2022 Moraes, Braz, Santos-Araujo, Oliveira, Bomfim, Ramos and Gondim.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures







References
-
- Alabaster A., Isoe J., Zhou G., Lee A., Murphy A., Day W. A., et al. (2011). Deficiencies in Acetyl-CoA Carboxylase and Fatty Acid Synthase 1 Differentially Affect Eggshell Formation and Blood Meal Digestion in Aedes aegypti . Insect Biochem. Mol. Biol. 41, 946–955. 10.1016/j.ibmb.2011.09.004 - DOI - PMC - PubMed
-
- Alves-Bezerra M., Ramos I. B., De Paula I. F., Maya-Monteiro C. M., Klett E. L., Coleman R. A., et al. (2017). Deficiency of Glycerol-3-Phosphate Acyltransferase 1 Decreases Triacylglycerol Storage and Induces Fatty Acid Oxidation in Insect Fat Body. Biochimica Biophysica Acta (BBA) - Mol. Cell Biol. Lipids 1862, 324–336. 10.1016/j.bbalip.2016.12.004 - DOI - PMC - PubMed
-
- Arêdes D. S., De Paula I. F., Santos-Araujo S., Gondim K. C. (2022). Silencing of Mitochondrial Trifunctional Protein A Subunit (HADHA) Increases Lipid Stores, and Reduces Oviposition and Flight Capacity in the Vector Insect Rhodnius prolixus . Front. Insect Sci. 2, 885172. 10.3389/finsc.2022.885172 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources