

How much does the unguarded X contribute to sex differences in life span?
- PMID: 35937469
- PMCID: PMC9346086
- DOI: 10.1002/evl3.292
How much does the unguarded X contribute to sex differences in life span?
Abstract
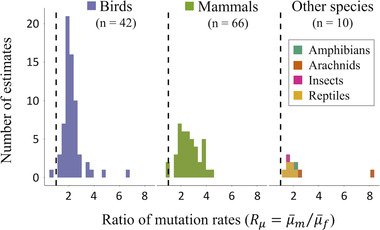
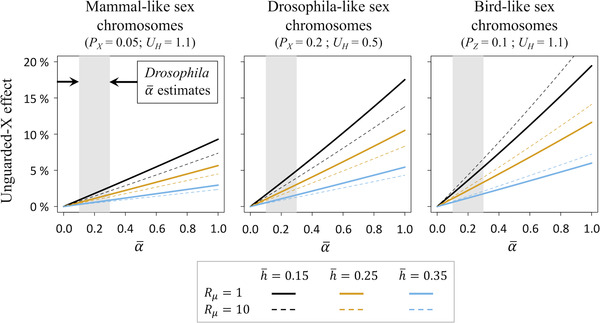
Females and males often have markedly different mortality rates and life spans, but it is unclear why these forms of sexual dimorphism evolve. The unguarded X hypothesis contends that dimorphic life spans arise from sex differences in X or Z chromosome copy number (i.e., one copy in the "heterogametic" sex; two copies in the "homogametic" sex), which leads to a disproportionate expression of deleterious mutations by the heterogametic sex (e.g., mammalian males; avian females). Although data on adult sex ratios and sex-specific longevity are consistent with predictions of the unguarded X hypothesis, direct experimental evidence remains scant, and alternative explanations are difficult to rule out. Using a simple population genetic model, we show that the unguarded X effect on sex differential mortality is a function of several reasonably well-studied evolutionary parameters, including the proportion of the genome that is sex linked, the genomic deleterious mutation rate, the mean dominance of deleterious mutations, the relative rates of mutation and strengths of selection in each sex, and the average effect of mutations on survival and longevity relative to their effects on fitness. We review published estimates of these parameters, parameterize our model with them, and show that unguarded X effects are too small to explain observed sex differences in life span across species. For example, sex differences in mean life span are known to often exceed 20% (e.g., in mammals), whereas our parameterized models predict unguarded X effects of a few percent (e.g., 1-3% in Drosophila and mammals). Indeed, these predicted unguarded X effects fall below statistical thresholds of detectability in most experiments, potentially explaining why direct tests of the hypothesis have generated little support for it. Our results suggest that evolution of sexually dimorphic life spans is predominantly attributable to other mechanisms, potentially including "toxic Y" effects and sexual dimorphism for optimal investment in survival versus reproduction.
Keywords: Deleterious mutations; evolution of life span; evolutionary theory; inbreeding depression; population genetics; sex chromosomes; sex ratio; sexual dimorphism.
© 2022 The Authors. Evolution Letters published by Wiley Periodicals LLC on behalf of Society for the Study of Evolution (SSE) and European Society for Evolutionary Biology (ESEB).
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Albert, A.Y.K. & Otto, S.P. (2005) Sexual selection can resolve sex‐linked sexual antagonism. Science, 310, 119–121. - PubMed
-
- Andrés, J.A. & Morrow, E.H. (2003) The origin of interlocus sexual conflict: is sex‐linkage important?. J. Evol. Biol, 16, 219–223. - PubMed
-
- Bonduriansky, R. , Maklakov, A. , Zajitschek, F. & Brooks, R. (2008) Sexual selection, sexual conflict and the evolution of ageing and life span. Funct. Ecol, 22, 443–453.
-
- Brengdahl, M. , Kimber, C.M. , Maguire‐Baxter, J. & Friberg, U. (2018) Sex differences in life span: females homozygous for the X chromosome do not suffer the shorter life span predicted by the unguarded X hypothesis. Evolution, 72, 568–577. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
