

Sex-specific natural selection on SNPs in Silene latifolia
- PMID: 35937470
- PMCID: PMC9346077
- DOI: 10.1002/evl3.283
Sex-specific natural selection on SNPs in Silene latifolia
Abstract
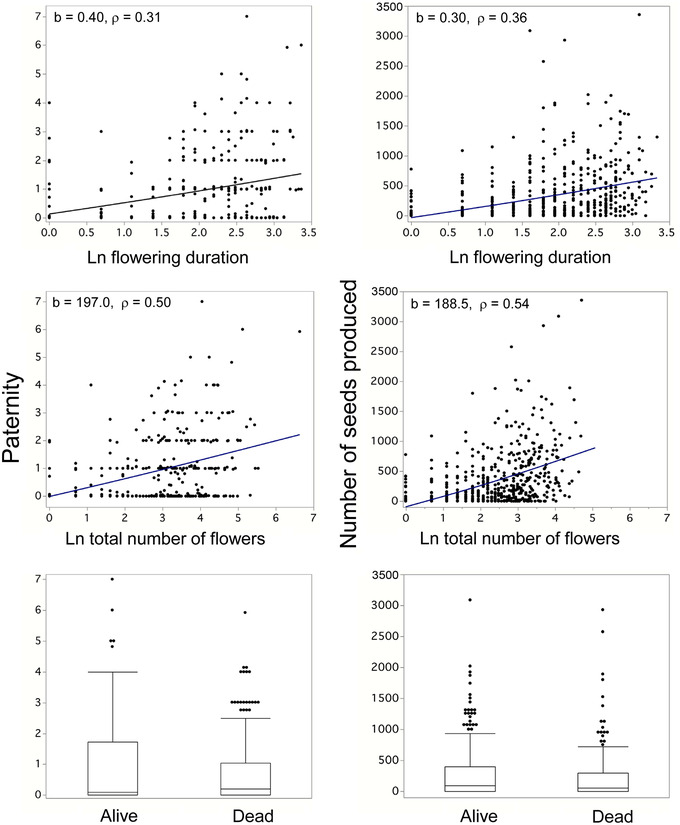
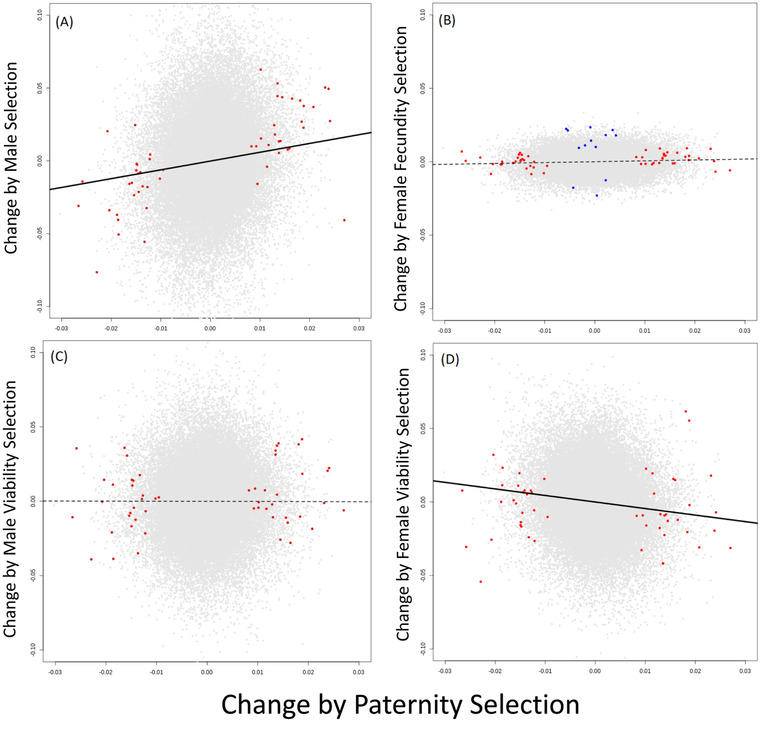
Selection that acts in a sex-specific manner causes the evolution of sexual dimorphism. Sex-specific phenotypic selection has been demonstrated in many taxa and can be in the same direction in the two sexes (differing only in magnitude), limited to one sex, or in opposing directions (antagonistic). Attempts to detect the signal of sex-specific selection from genomic data have confronted numerous difficulties. These challenges highlight the utility of "direct approaches," in which fitness is predicted from individual genotype within each sex. Here, we directly measured selection on Single Nucleotide Polymorphisms (SNPs) in a natural population of the sexually dimorphic, dioecious plant, Silene latifolia. We measured flowering phenotypes, estimated fitness over one reproductive season, as well as survival to the next year, and genotyped all adults and a subset of their offspring for SNPs across the genome. We found that while phenotypic selection was congruent (fitness covaried similarly with flowering traits in both sexes), SNPs showed clear evidence for sex-specific selection. SNP-level selection was particularly strong in males and may involve an important gametic component (e.g., pollen competition). While the most significant SNPs under selection in males differed from those under selection in females, paternity selection showed a highly polygenic tradeoff with female survival. Alleles that increased male mating success tended to reduce female survival, indicating sexual antagonism at the genomic level. Perhaps most importantly, this experiment demonstrates that selection within natural populations can be strong enough to measure sex-specific fitness effects of individual loci. Males and females typically differ phenotypically, a phenomenon known as sexual dimorphism. These differences arise when selection on males differs from selection on females, either in magnitude or direction. Estimated relationships between traits and fitness indicate that sex-specific selection is widespread, occurring in both plants and animals, and explains why so many species exhibit sexual dimorphism. Finding the specific loci experiencing sex-specific selection is a challenging prospect but one worth undertaking given the extensive evolutionary consequences. Flowering plants with separate sexes are ideal organisms for such studies, given that the fitness of females can be estimated by counting the number of seeds they produce. Determination of fitness for males has been made easier as thousands of genetic markers can now be used to assign paternity to seeds. We undertook just such a study in S. latifolia, a short-lived, herbaceous plant. We identified loci under sex-specific selection in this species and found more loci affecting fitness in males than females. Importantly, loci with major effects on male fitness were distinct from the loci with major effects on females. We detected sexual antagonism only when considering the aggregate effect of many loci. Hence, even though males and females share the same genome, this does not necessarily impose a constraint on their independent evolution.
Keywords: dioecious; fitness; paternity; selection component analysis; sexual dimorphism; sex‐specific selection.
© 2022 The Authors. Evolution Letters published by Wiley Periodicals LLC on behalf of Society for the Study of Evolution (SSE) and European Society for Evolutionary Biology (ESEB).
Figures

Similar articles
-
Temporal dynamics of sexual dimorphism in a dioecious species.Ann Bot. 2020 Aug 13;126(3):471-480. doi: 10.1093/aob/mcaa088. Ann Bot. 2020. PMID: 32386315 Free PMC article.
-
The genomic architecture of sexual dimorphism in the dioecious plant Silene latifolia.Evolution. 2010 Oct;64(10):2873-86. doi: 10.1111/j.1558-5646.2010.01048.x. Epub 2010 Aug 19. Evolution. 2010. PMID: 20550575
-
Fungal Infection Induces Sex-Specific Transcriptional Changes and Alters Sexual Dimorphism in the Dioecious Plant Silene latifolia.PLoS Genet. 2015 Oct 8;11(10):e1005536. doi: 10.1371/journal.pgen.1005536. eCollection 2015 Oct. PLoS Genet. 2015. PMID: 26448481 Free PMC article.
-
An integrative view of sexual selection in Tribolium flour beetles.Biol Rev Camb Philos Soc. 2008 May;83(2):151-71. doi: 10.1111/j.1469-185X.2008.00037.x. Biol Rev Camb Philos Soc. 2008. PMID: 18429767 Review.
-
Sexual conflict and speciation.Philos Trans R Soc Lond B Biol Sci. 1998 Feb 28;353(1366):261-74. doi: 10.1098/rstb.1998.0208. Philos Trans R Soc Lond B Biol Sci. 1998. PMID: 9533125 Free PMC article. Review.
Cited by
-
The battle of the sexes in humans is highly polygenic.Proc Natl Acad Sci U S A. 2024 Sep 24;121(39):e2412315121. doi: 10.1073/pnas.2412315121. Epub 2024 Sep 20. Proc Natl Acad Sci U S A. 2024. PMID: 39302970 Free PMC article.
-
Costs of reproduction in flowering plants.New Phytol. 2025 Jul;247(1):55-70. doi: 10.1111/nph.70166. Epub 2025 May 7. New Phytol. 2025. PMID: 40342244 Free PMC article. Review.
-
Defining fitness in evolutionary ecology.Int J Plant Sci. 2024 May;185(3):218-227. doi: 10.1086/729360. Epub 2024 Apr 19. Int J Plant Sci. 2024. PMID: 39035046 Free PMC article.
-
Taking quantitative genomics into the wild.Proc Biol Sci. 2022 Dec 21;289(1989):20221930. doi: 10.1098/rspb.2022.1930. Epub 2022 Dec 21. Proc Biol Sci. 2022. PMID: 36541172 Free PMC article.
-
Two Forms of Sexual Dimorphism in Gene Expression in Drosophila melanogaster: Their Coincidence and Evolutionary Genetics.Mol Biol Evol. 2023 May 2;40(5):msad091. doi: 10.1093/molbev/msad091. Mol Biol Evol. 2023. PMID: 37116199 Free PMC article.
References
-
- Allen, S.L. , McGuigan, K. , Connallon, T. , Blows, M.W. & Chenoweth, S.F. (2017) Sexual selection on spontaneous mutations strengthens the between‐sex genetic correlation for fitness. Evolution; Internation Journal of Organic Evolution, 71, 2398–2409. - PubMed
-
- Arnqvist, G. & Rowe, L. (2005) Sexual conflict. Princeton, NJ: Princeton University Press.
-
- Arunkumar, R. , Josephs, E.B. , Williamson, R.J. & Wright, S.I. (2013) Pollen‐specific, but not sperm‐specific, genes show stronger purifying selection and higher rates of positive selection than sporophytic genes in Capsella grandiflora . Mol. Biol. Evol,. 30, 2475–2486. - PubMed
