

The Metabolic Adaptation in Response to Nitrate Is Critical for Actinobacillus pleuropneumoniae Growth and Pathogenicity under the Regulation of NarQ/P
- PMID: 35938858
- PMCID: PMC9476948
- DOI: 10.1128/iai.00239-22
The Metabolic Adaptation in Response to Nitrate Is Critical for Actinobacillus pleuropneumoniae Growth and Pathogenicity under the Regulation of NarQ/P
Abstract
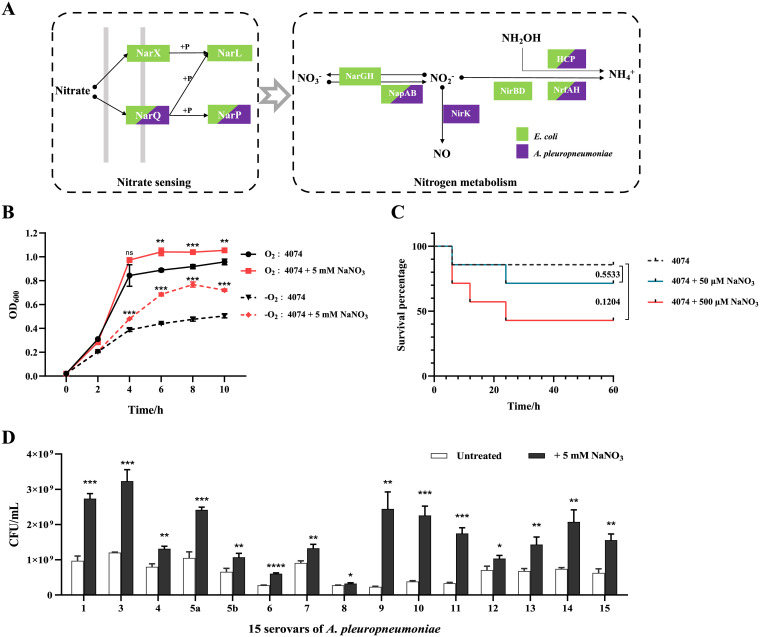
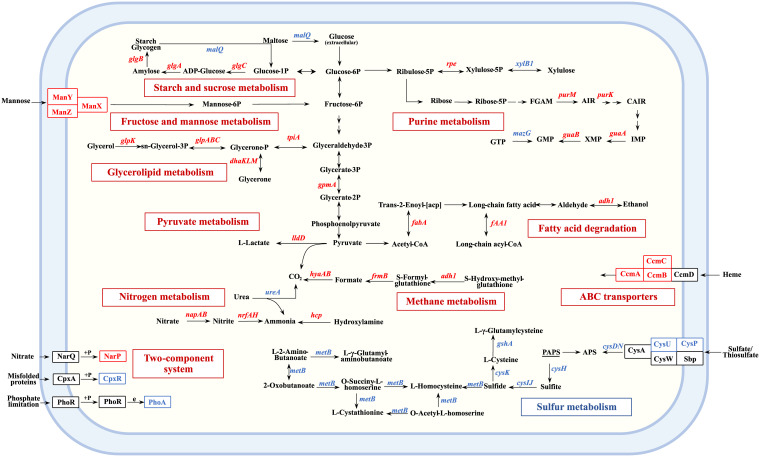
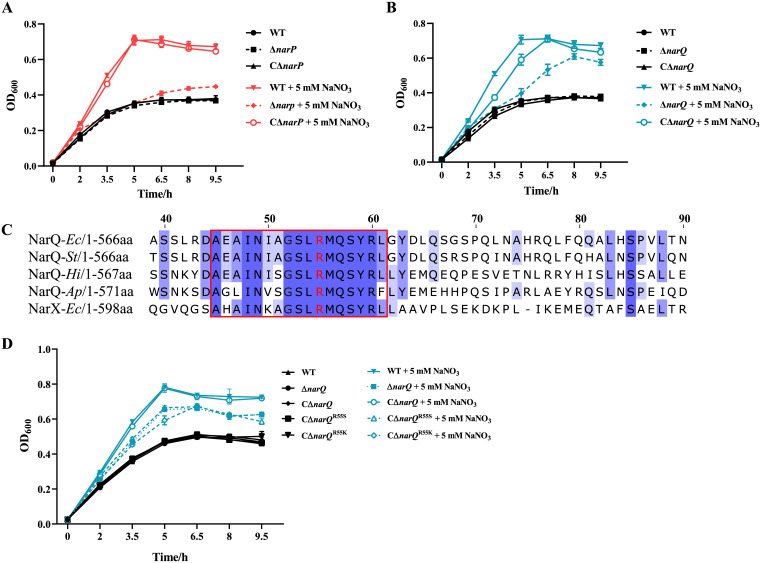
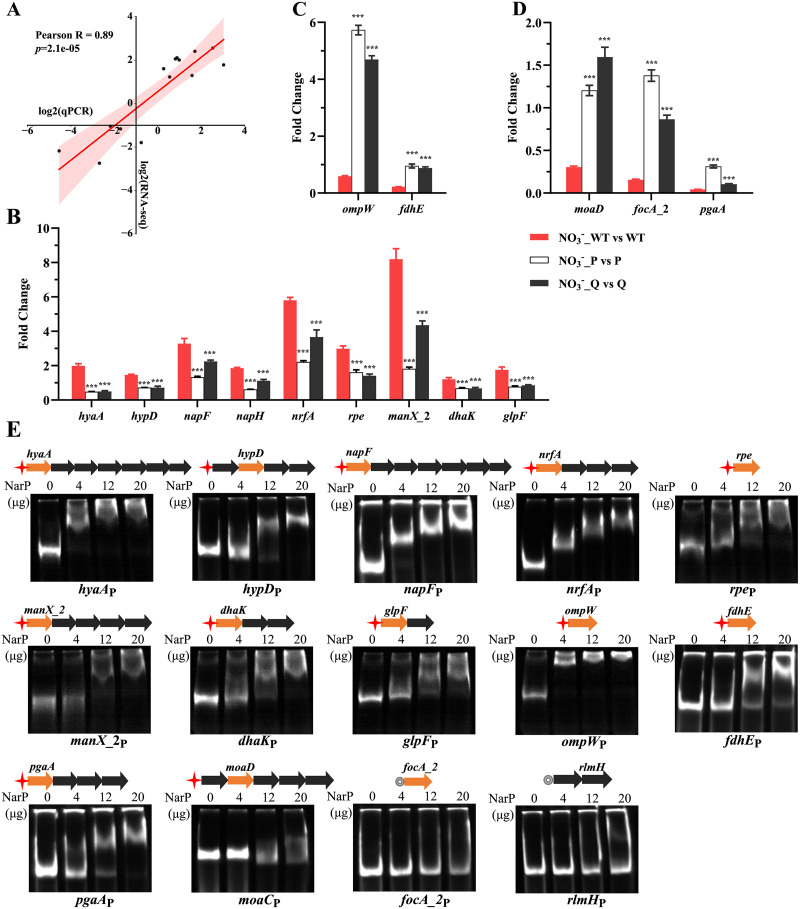
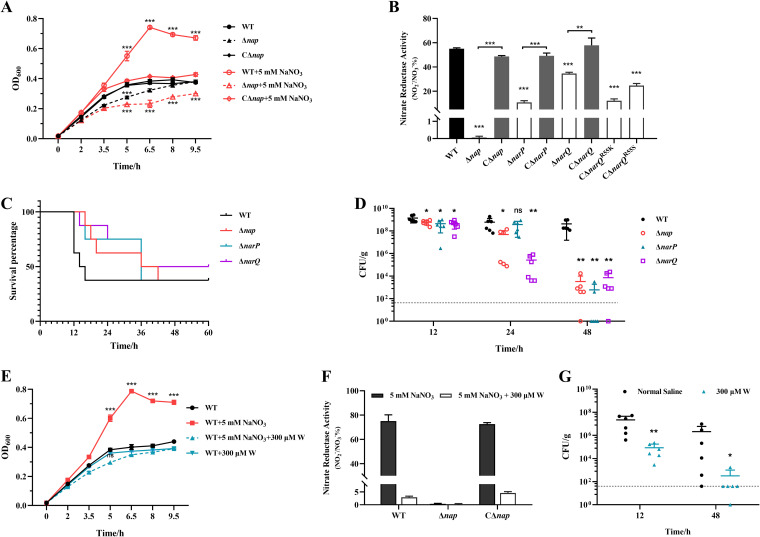
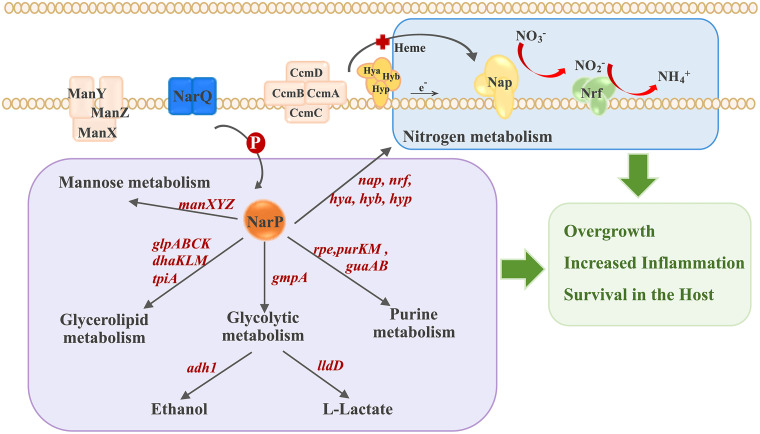
Nitrate metabolism is an adaptation mechanism used by many bacteria for survival in anaerobic environments. As a by-product of inflammation, nitrate is used by the intestinal bacterial pathogens to enable gut infection. However, the responses of bacterial respiratory pathogens to nitrate are less well understood. Actinobacillus pleuropneumoniae is an important bacterial respiratory pathogen of swine. Previous studies have suggested that adaptation of A. pleuropneumoniae to anaerobiosis is important for infection. In this work, A. pleuropneumoniae growth and pathogenesis in response to the nitrate were investigated. Nitrate significantly promoted A. pleuropneumoniae growth under anaerobic conditions in vitro and lethality in mice. By using narQ and narP deletion mutants and single-residue-mutated complementary strains of ΔnarQ, the two-component system NarQ/P was confirmed to be critical for nitrate-induced growth, with Arg50 in NarQ as an essential functional residue. Transcriptome analysis showed that nitrate upregulated multiple energy-generating pathways, including nitrate metabolism, mannose and pentose metabolism, and glycerolipid metabolism via the regulation of NarQ/P. Furthermore, narQ, narP, and its target gene encoding the nitrate reductase Nap contributed to the pathogenicity of A. pleuropneumoniae. The Nap inhibitor tungstate significantly reduced the survival of A. pleuropneumoniae in vivo, suggesting that Nap is a potential drug target. These results give new insights into how the respiratory pathogen A. pleuropneumoniae utilizes the alternative electron acceptor nitrate to overcome the hypoxia microenvironment, which can occur in the inflammatory or necrotic infected tissues.
Keywords: Actinobacillus pleuropneumoniae; NarP; NarQ; growth; nitrate; pathogenicity; regulation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Identification of genes regulated by the two-component system response regulator NarP of Actinobacillus pleuropneumoniae via DNA-affinity-purified sequencing.Microbiol Res. 2020 Jan;230:126343. doi: 10.1016/j.micres.2019.126343. Epub 2019 Sep 10. Microbiol Res. 2020. PMID: 31539852
-
'Locked-on' and 'locked-off' signal transduction mutations in the periplasmic domain of the Escherichia coli NarQ and NarX sensors affect nitrate- and nitrite-dependent regulation by NarL and NarP.Mol Microbiol. 1997 Jun;24(5):1049-60. doi: 10.1046/j.1365-2958.1997.4131779.x. Mol Microbiol. 1997. PMID: 9220011
-
Identification of FtpA, a Dps-Like Protein Involved in Anti-Oxidative Stress and Virulence in Actinobacillus pleuropneumoniae.J Bacteriol. 2022 Feb 15;204(2):e0032621. doi: 10.1128/JB.00326-21. Epub 2021 Nov 22. J Bacteriol. 2022. PMID: 34807725 Free PMC article.
-
Actinobacillus pleuropneumoniae biofilms: Role in pathogenicity and potential impact for vaccination development.Anim Health Res Rev. 2018 Jun;19(1):17-30. doi: 10.1017/S146625231700010X. Epub 2017 Nov 7. Anim Health Res Rev. 2018. PMID: 29110751 Review.
-
Virulence factors of the swine pathogen Actinobacillus pleuropneumoniae.Microbiologia. 1996 Jun;12(2):171-84. Microbiologia. 1996. PMID: 8767702 Review.
Cited by
-
Transcriptional Response of Pectobacterium carotovorum to Cinnamaldehyde Treatment.J Microbiol Biotechnol. 2024 Mar 28;34(3):538-546. doi: 10.4014/jmb.2311.11043. Epub 2023 Dec 25. J Microbiol Biotechnol. 2024. PMID: 38146216 Free PMC article.
-
HemN2 Regulates the Virulence of Pseudomonas donghuensis HYS through 7-Hydroxytropolone Synthesis and Oxidative Stress.Biology (Basel). 2024 May 24;13(6):373. doi: 10.3390/biology13060373. Biology (Basel). 2024. PMID: 38927253 Free PMC article.
-
Development and Characterization of a Recombinant galT-galU Protein for Broad-Spectrum Immunoprotection Against Porcine Contagious Pleuropneumonia.Int J Mol Sci. 2025 Apr 11;26(8):3634. doi: 10.3390/ijms26083634. Int J Mol Sci. 2025. PMID: 40332240 Free PMC article.
-
[Effect of intestinal nitrate on growth of Klebsiella pneumoniae and its regulatory mechanism].Nan Fang Yi Ke Da Xue Xue Bao. 2024 Apr 20;44(4):757-764. doi: 10.12122/j.issn.1673-4254.2024.04.18. Nan Fang Yi Ke Da Xue Xue Bao. 2024. PMID: 38708510 Free PMC article. Chinese.
-
The morphology and metabolic changes of Actinobacillus pleuropneumoniae during its growth as a biofilm.Vet Res. 2023 May 26;54(1):42. doi: 10.1186/s13567-023-01173-x. Vet Res. 2023. PMID: 37237397 Free PMC article.
References
-
- Winter SE, Winter MG, Xavier MN, Thiennimitr P, Poon V, Keestra AM, Laughlin RC, Gomez G, Wu J, Lawhon SD, Popova IE, Parikh SJ, Adams LG, Tsolis RM, Stewart VJ, Bäumler AJ. 2013. Host-derived nitrate boosts growth of E. coli in the inflamed gut. Science 339:708–711. 10.1126/science.1232467. - DOI - PMC - PubMed
-
- Stecher B, Robbiani R, Walker AW, Westendorf AM, Barthel M, Kremer M, Chaffron S, Macpherson AJ, Buer J, Parkhill J, Dougan G, von Mering C, Hardt WD. 2007. Salmonella enterica serovar Typhimurium exploits inflammation to compete with the intestinal microbiota. PLoS Biol 5:2177–2189. 10.1371/journal.pbio.0050244. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
