

Insight into Viral Hijacking of CRL4 Ubiquitin Ligase through Structural Analysis of the pUL145-DDB1 Complex
- PMID: 35938868
- PMCID: PMC9472758
- DOI: 10.1128/jvi.00826-22
Insight into Viral Hijacking of CRL4 Ubiquitin Ligase through Structural Analysis of the pUL145-DDB1 Complex
Abstract
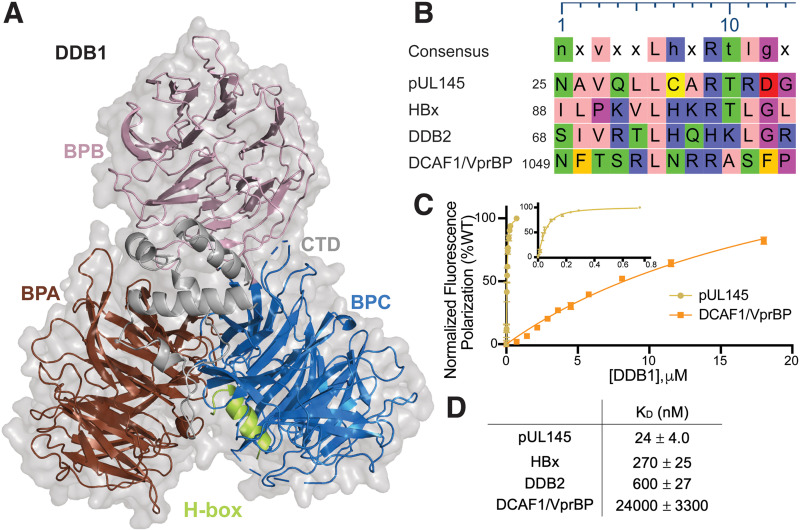
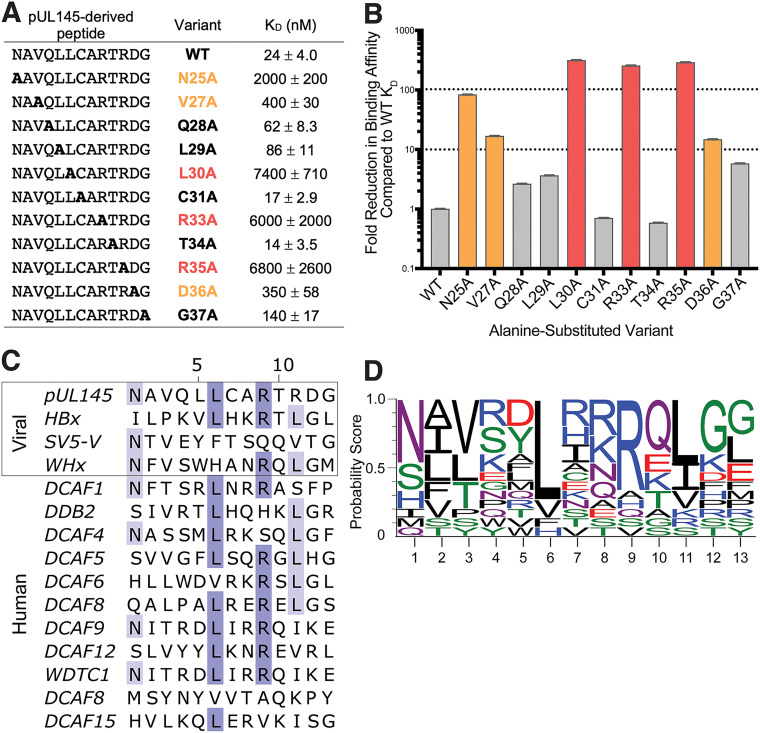
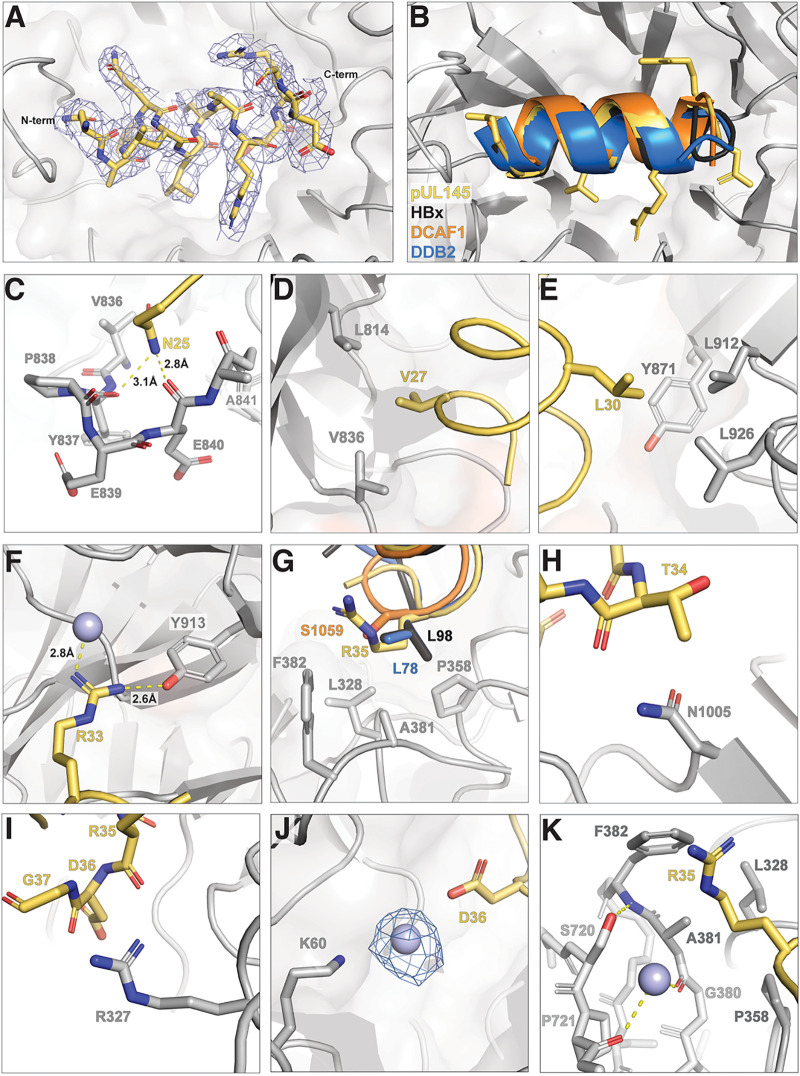
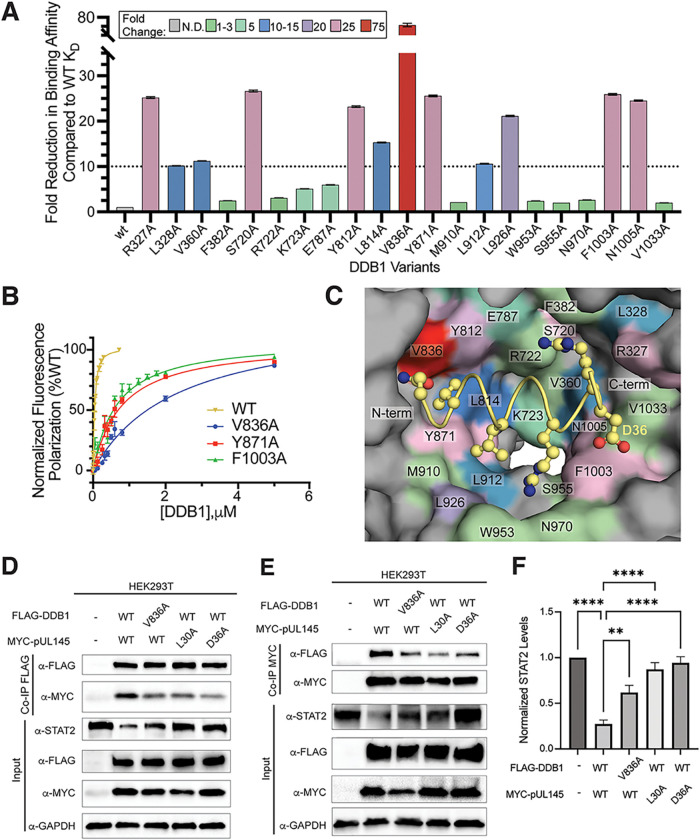
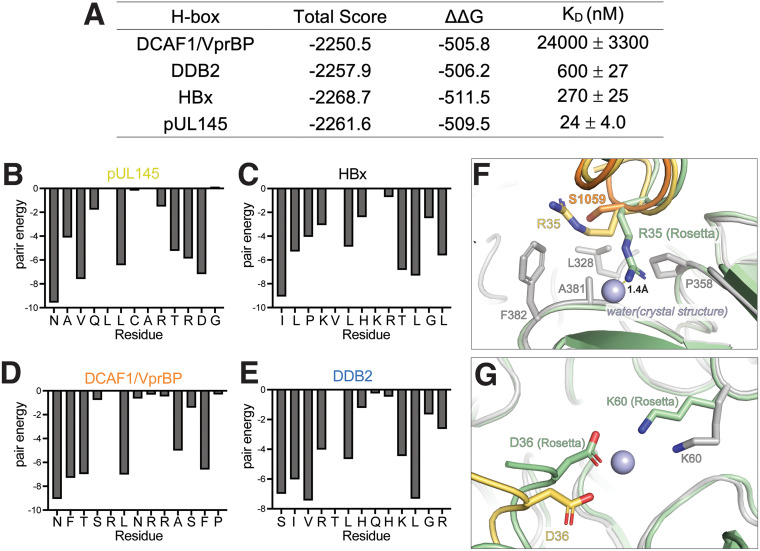
Viruses evolve mechanisms to exploit cellular pathways that increase viral fitness, e.g., enhance viral replication or evade the host cell immune response. The ubiquitin-proteosome system, a fundamental pathway-regulating protein fate in eukaryotes, is hijacked by all seven classes of viruses. Members of the Cullin-RING family of ubiquitin (Ub) ligases are frequently co-opted by divergent viruses because they can target a broad array of substrates by forming multisubunit assemblies comprised of a variety of adapters and substrate receptors. For example, the linker subunit DDB1 in the cullin 4-RING (CRL4)-DDB1 Ub ligase (CRL4DDB1) interacts with an H-box motif found in several unrelated viral proteins, including the V protein of simian virus 5 (SV5-V), the HBx protein of hepatitis B virus (HBV), and the recently identified pUL145 protein of human cytomegalovirus (HCMV). In HCMV-infected cells, pUL145 repurposes CRL4DDB1 to target STAT2, a protein vital to the antiviral immune response. However, the details of how these divergent viral sequences hijack DDB1 is not well understood. Here, we use a combination of binding assays, X-ray crystallography, alanine scanning, cell-based assays, and computational analysis to reveal that viral H-box motifs appear to bind to DDB1 with a higher affinity than the H-box motifs from host proteins DCAF1 and DDB2. This analysis reveals that viruses maintain native hot-spot residues in the H-box motif of host DCAFs and also acquire favorable interactions at neighboring residues within the H-box. Overall, these studies reveal how viruses evolve strategies to produce high-affinity binding and quality interactions with DDB1 to repurpose its Ub ligase machinery. IMPORTANCE Many different viruses modulate the protein machinery required for ubiquitination to enhance viral fitness. Specifically, several viruses hijack the cullin-RING ligase CRL4DDB1 to degrade host resistance factors. Human cytomegalovirus (HCMV) encodes pUL145 that redirects CRL4DDB1 to evade the immune system through the targeted degradation of the antiviral immune response protein STAT2. However, it is unclear why several viruses bind specific surfaces on ubiquitin ligases to repurpose their activity. We demonstrate that viruses have optimized H-box motifs that bind DDB1 with higher affinity than the H-box of native binders. For viral H-boxes, native interactions are maintained, but additional interactions that are absent in host cell H-boxes are formed, indicating that rewiring CRL4DDB1 creates a selective advantage for the virus. The DDB1-pUL145 peptide structure reveals that water-mediated interactions are critical to the higher affinity. Together, our data present an interesting example of how viral evolution can exploit a weakness in the ubiquitination machinery.
Keywords: cullin-RING; human cytomegalovirus; innate immunity; ubiquitin ligase; viral hijacking.
Conflict of interest statement
The authors declare a conflict of interest. Y.X. is an inventor named on patent applications filed by UNC. The Xiong laboratory has received research funds from Cullgen, Inc. Y.X. is an equity shareholder in and employee of Cullgen, Inc. All other authors declare no competing interesting.
Figures
Similar articles
-
The Human Cytomegalovirus pUL145 Isoforms Act as Viral DDB1-Cullin-Associated Factors to Instruct Host Protein Degradation to Impede Innate Immunity.Cell Rep. 2020 Feb 18;30(7):2248-2260.e5. doi: 10.1016/j.celrep.2020.01.070. Cell Rep. 2020. PMID: 32075763
-
Hepatitis B Virus HBx Protein Mediates the Degradation of Host Restriction Factors through the Cullin 4 DDB1 E3 Ubiquitin Ligase Complex.Cells. 2020 Mar 30;9(4):834. doi: 10.3390/cells9040834. Cells. 2020. PMID: 32235678 Free PMC article.
-
A Mass Spectrometry-Based Profiling of Interactomes of Viral DDB1- and Cullin Ubiquitin Ligase-Binding Proteins Reveals NF-κB Inhibitory Activity of the HIV-2-Encoded Vpx.Front Immunol. 2018 Dec 19;9:2978. doi: 10.3389/fimmu.2018.02978. eCollection 2018. Front Immunol. 2018. PMID: 30619335 Free PMC article.
-
CRL4s: the CUL4-RING E3 ubiquitin ligases.Trends Biochem Sci. 2009 Nov;34(11):562-70. doi: 10.1016/j.tibs.2009.07.002. Epub 2009 Oct 7. Trends Biochem Sci. 2009. PMID: 19818632 Free PMC article. Review.
-
Cullin E3 ligases and their rewiring by viral factors.Biomolecules. 2014 Oct 13;4(4):897-930. doi: 10.3390/biom4040897. Biomolecules. 2014. PMID: 25314029 Free PMC article. Review.
Cited by
-
Structural mechanism of CRL4-instructed STAT2 degradation via a novel cytomegaloviral DCAF receptor.EMBO J. 2023 Mar 1;42(5):e112351. doi: 10.15252/embj.2022112351. Epub 2023 Feb 10. EMBO J. 2023. PMID: 36762436 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
