

IgM antibodies derived from memory B cells are potent cross-variant neutralizers of SARS-CoV-2
- PMID: 35938988
- PMCID: PMC9365875
- DOI: 10.1084/jem.20220849
IgM antibodies derived from memory B cells are potent cross-variant neutralizers of SARS-CoV-2
Erratum in
-
Correction: IgM antibodies derived from memory B cells are potent cross-variant neutralizers of SARS-CoV-2.J Exp Med. 2022 Sep 5;219(9):jem.2022084908172022c. doi: 10.1084/jem.2022084908172022c. Epub 2022 Aug 29. J Exp Med. 2022. PMID: 36036783 Free PMC article. No abstract available.
Abstract
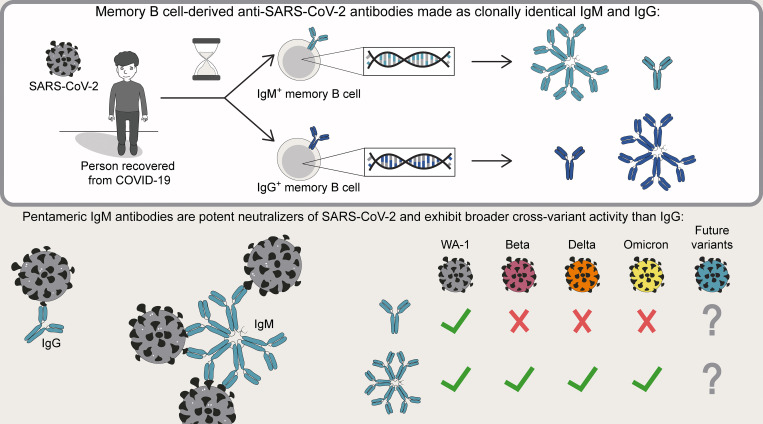
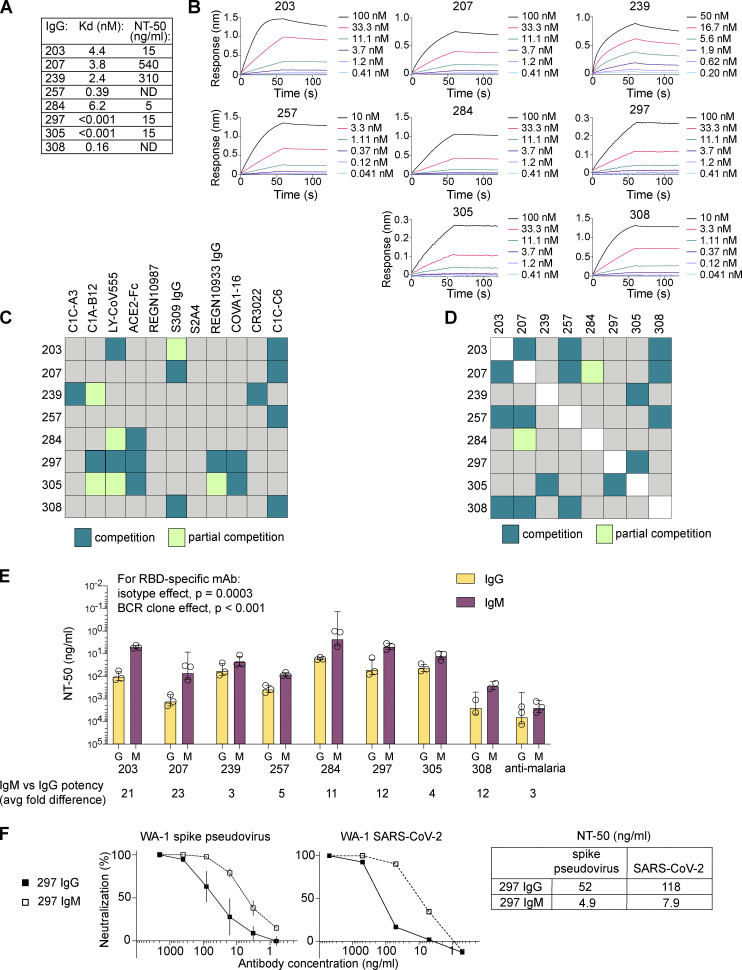
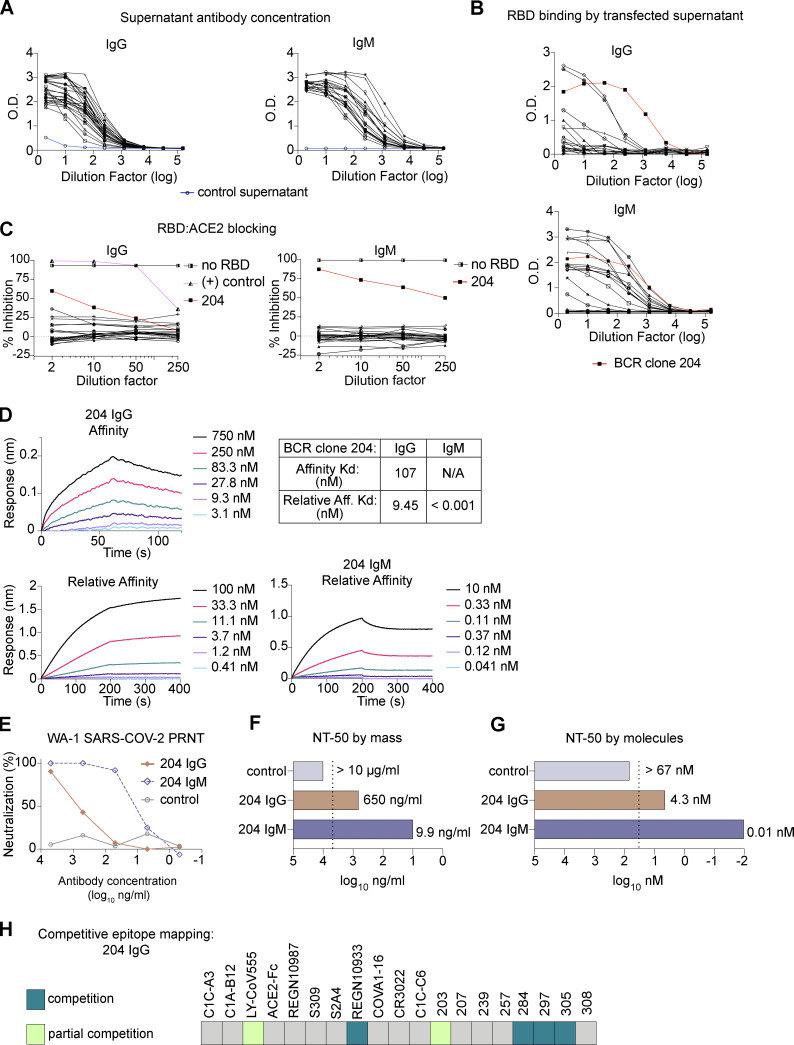
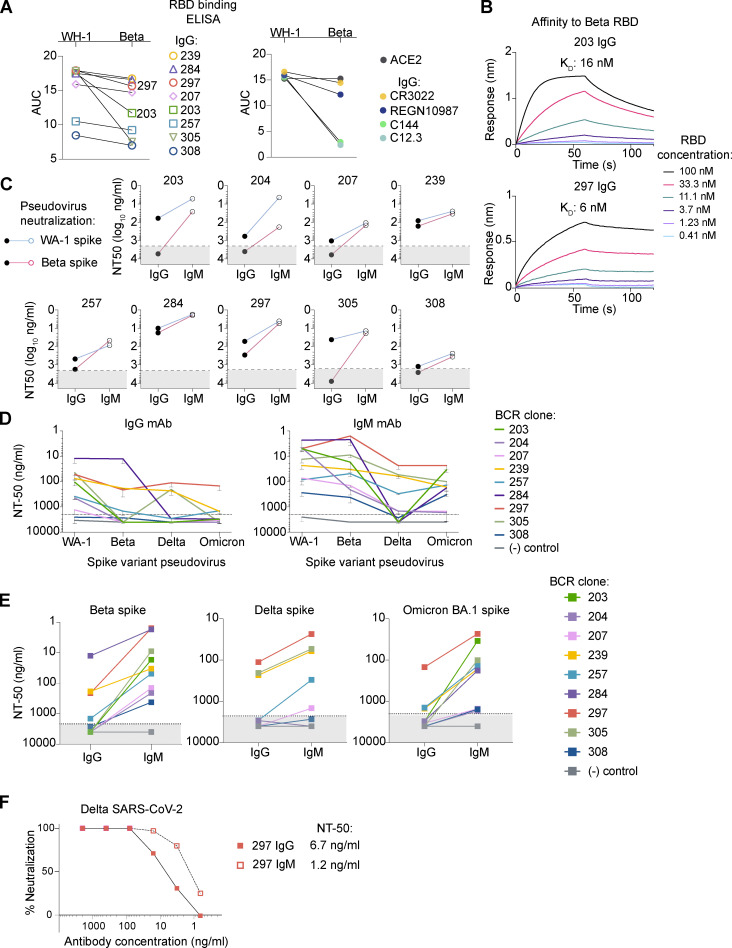
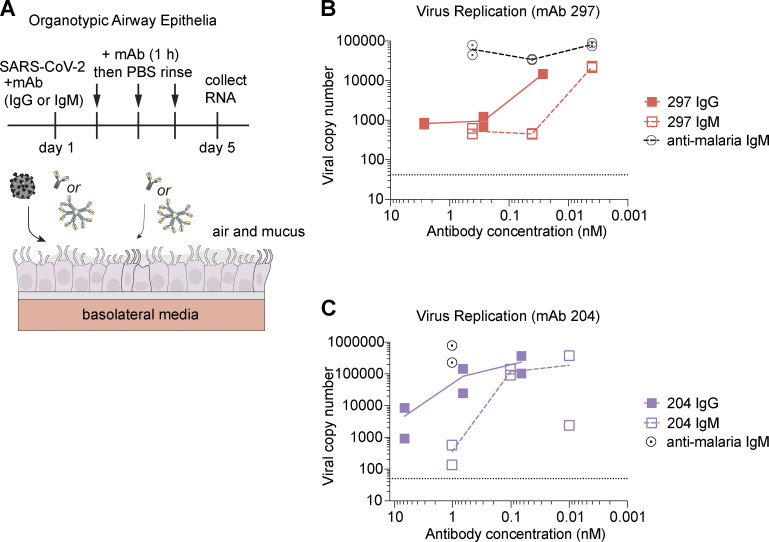
Humoral immunity to SARS-CoV-2 can be supplemented with polyclonal sera from convalescent donors or an engineered monoclonal antibody (mAb) product. While pentameric IgM antibodies are responsible for much of convalescent sera's neutralizing capacity, all available mAbs are based on the monomeric IgG antibody subtype. We now show that IgM mAbs derived from immune memory B cell receptors are potent neutralizers of SARS-CoV-2. IgM mAbs outperformed clonally identical IgG antibodies across a range of affinities and SARS-CoV-2 receptor-binding domain epitopes. Strikingly, efficacy against SARS-CoV-2 viral variants was retained for IgM but not for clonally identical IgG. To investigate the biological role for IgM memory in SARS-CoV-2, we also generated IgM mAbs from antigen-experienced IgM+ memory B cells in convalescent donors, identifying a potent neutralizing antibody. Our results highlight the therapeutic potential of IgM mAbs and inform our understanding of the role for IgM memory against a rapidly mutating pathogen.
© 2022 Hale et al.
Conflict of interest statement
Disclosures: M. Hale, J. Netland, Y. Chen, C.D. Thouvenel, L.B. Rodda, M. Pepper, and D.J. Rawlings report a patent to SARS-CoV-2–neutralizing antibodies, biomarkers to predict protection from re-infection, and high efficiency antibody screening methods, PCT/US2021/045427 pending. N.P. King is a co-founder, shareholder, paid consultant, and chair of the scientific advisory board of Icosavax, Inc. The King lab has received unrelated sponsored research agreements from Pfizer and GSK. M. Pepper reported personal fees from the Vaxart Scientific Advisory Board and the Neoleukin Scientific Advisory Board outside the submitted work. No other disclosures were reported.
Figures


References
-
- Alamyar, E., Duroux P., Lefranc M.-P., and Giudicelli V.. 2012. IMGT(®) tools for the nucleotide analysis of immunoglobulin (IG) and T cell receptor (TR) V-(D)-J repertoires, polymorphisms, and IG mutations: IMGT/V-QUEST and IMGT/HighV-QUEST for NGS. Methods Mol. Biol. 882:569–604. 10.1007/978-1-61779-842-9_32 - DOI - PubMed
-
- Altman, M.C., Reeves S.R., Parker A.R., Whalen E., Misura K.M., Barrow K.A., James R.G., Hallstrand T.S., Ziegler S.F., and Debley J.S.. 2018. Interferon response to respiratory syncytial virus by bronchial epithelium from children with asthma is inversely correlated with pulmonary function. J. Allergy Clin. Immunol. 142:451–459. 10.1016/j.jaci.2017.10.004 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
- T32 AI106677/AI/NIAID NIH HHS/United States
- S10 OD024979/OD/NIH HHS/United States
- T32 AI007140/AI/NIAID NIH HHS/United States
- T32 GM007753/GM/NIGMS NIH HHS/United States
- T32 GM144273/GM/NIGMS NIH HHS/United States
- U01 AI151698/AI/NIAID NIH HHS/United States
- Burroughs Wellcome Fund
- U01 AI142001/AI/NIAID NIH HHS/United States
- K24 AI150991/AI/NIAID NIH HHS/United States
- OPP1156262/GATES/Bill & Melinda Gates Foundation/United States
- R01 AI163160/AI/NIAID NIH HHS/United States
- R01 AI153191/AI/NIAID NIH HHS/United States
- T32 GM007266/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
