

Noradrenaline activation of hippocampal dopamine D1 receptors promotes antidepressant effects
- PMID: 35939697
- PMCID: PMC9388128
- DOI: 10.1073/pnas.2117903119
Noradrenaline activation of hippocampal dopamine D1 receptors promotes antidepressant effects
Abstract
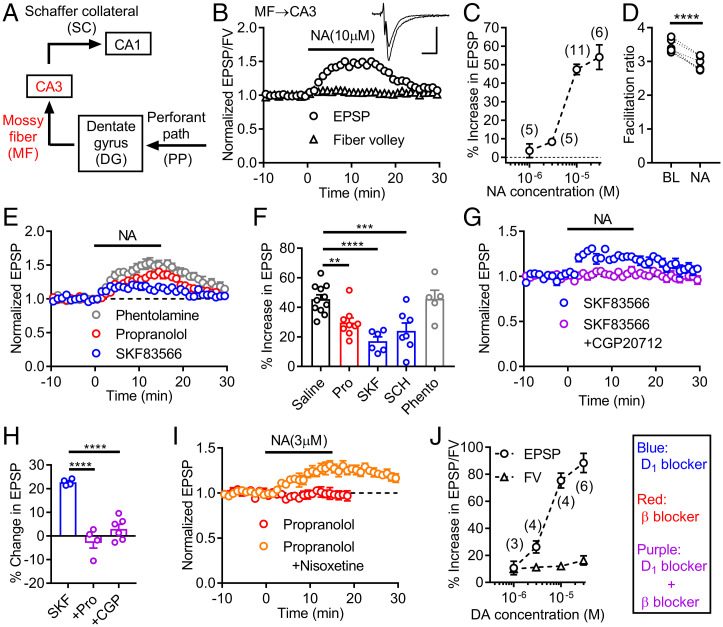
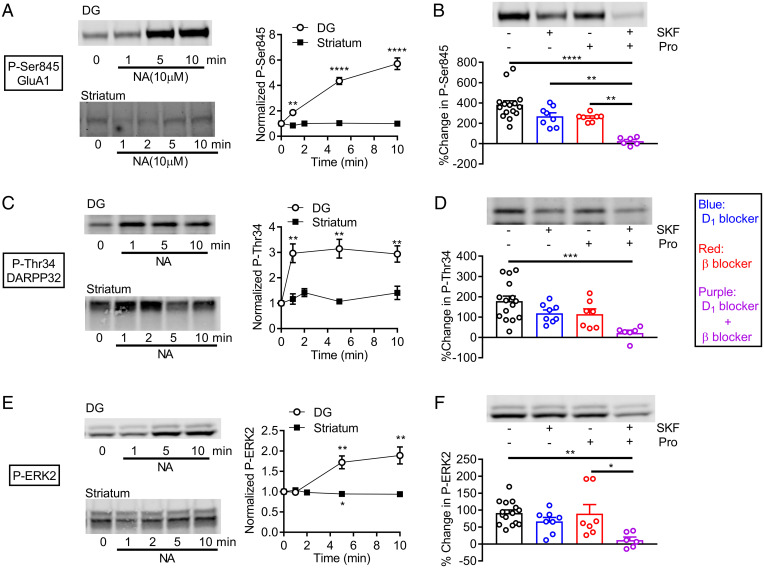
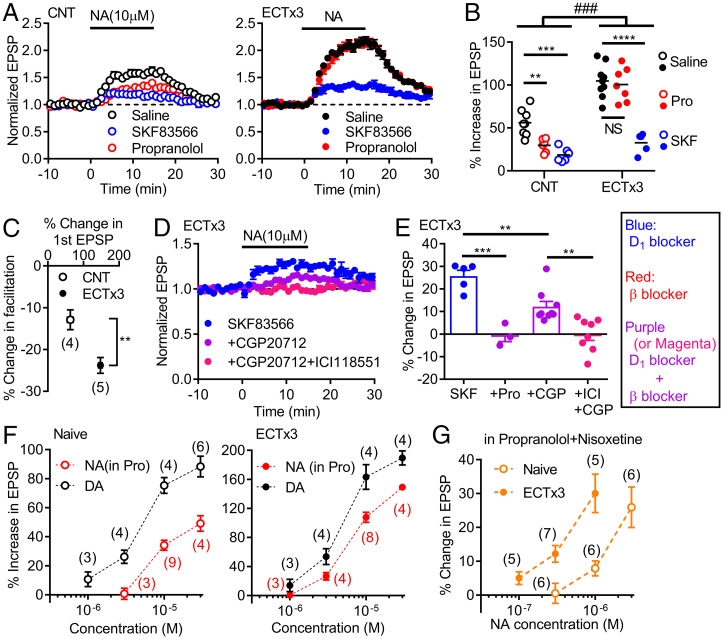
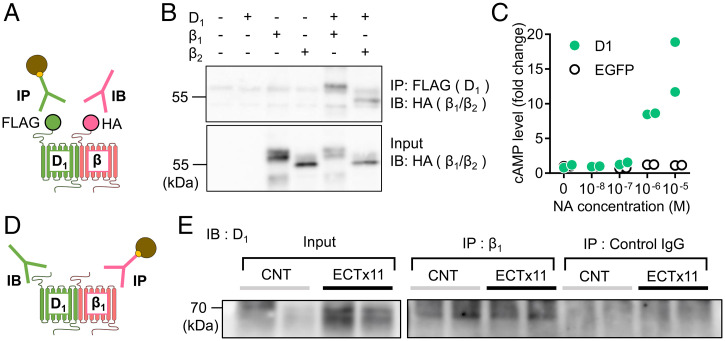
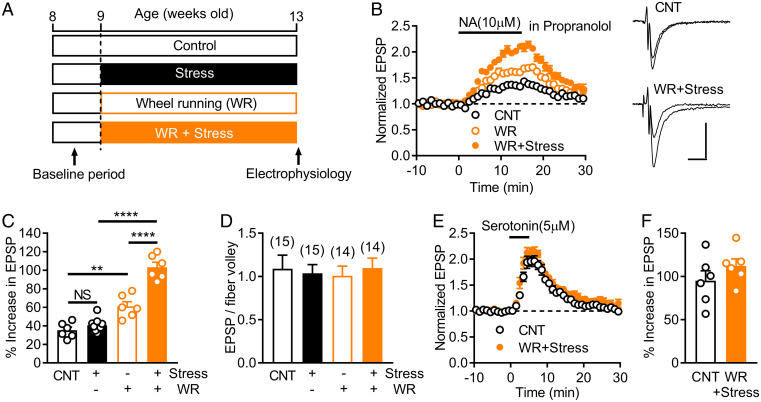
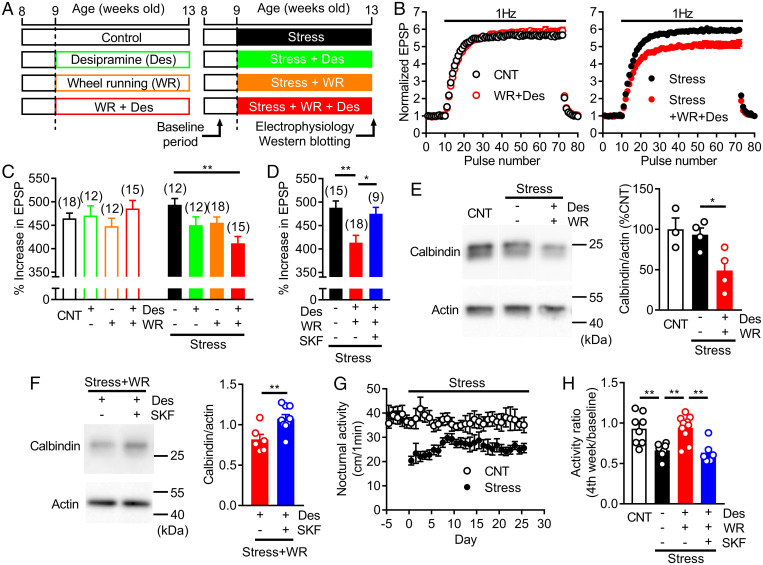
Dopamine D1 receptors (D1Rs) in the hippocampal dentate gyrus (DG) are essential for antidepressant effects. However, the midbrain dopaminergic neurons, the major source of dopamine in the brain, only sparsely project to DG, suggesting possible activation of DG D1Rs by endogenous substances other than dopamine. We have examined this possibility using electrophysiological and biochemical techniques and found robust activation of D1Rs in mouse DG neurons by noradrenaline. Noradrenaline at the micromolar range potentiated synaptic transmission at the DG output and increased the phosphorylation of protein kinase A substrates in DG via activation of D1Rs and β adrenergic receptors. Neuronal excitation preferentially enhanced noradrenaline-induced synaptic potentiation mediated by D1Rs with minor effects on β-receptor-dependent potentiation. Increased voluntary exercise by wheel running also enhanced noradrenaline-induced, D1R-mediated synaptic potentiation, suggesting a distinct functional role of the noradrenaline-D1R signaling. We then examined the role of this signaling in antidepressant effects using mice exposed to chronic restraint stress. In the stressed mice, an antidepressant acting on the noradrenergic system induced a mature-to-immature change in the DG neuron phenotype, a previously proposed cellular substrate for antidepressant action. This effect was evident only in mice subjected to wheel running and blocked by a D1R antagonist. These results suggest a critical role of noradrenaline-induced activation of D1Rs in antidepressant effects in DG. Experience-dependent regulation of noradrenaline-D1R signaling may determine responsiveness to antidepressant drugs in depressive disorders.
Keywords: dentate gyrus; exercise; monoamine; mossy fiber; stress.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- da Silva W. C., Köhler C. C., Radiske A., Cammarota M., D1/D5 dopamine receptors modulate spatial memory formation. Neurobiol. Learn. Mem. 97, 271–275 (2012). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
