

Review
doi: 10.1101/cshperspect.a041258.
Sorting and Export of Proteins at the Endoplasmic Reticulum
Affiliations
- PMID: 35940902
- PMCID: PMC10153803
- DOI: 10.1101/cshperspect.a041258
Item in Clipboard
Review
Sorting and Export of Proteins at the Endoplasmic Reticulum
Cold Spring Harb Perspect Biol.
.
Abstract
Secretory proteins are transported from the endoplasmic reticulum (ER) to the Golgi complex in carriers that are formed by the concerted activities of cytoplasmic proteins in the coat protein complex II (COPII). COPII was first described in Saccharomyces cerevisiae and its basic functions are largely conserved throughout eukaryotes. The discovery of the TANGO1 (transport and Golgi organization 1) family of proteins is revealing insights into how cells can adapt COPII proteins to reorganize the ER exit site for the export of the most abundant and bulky molecules, collagens.
Copyright © 2023 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures

The secretory pathway. A schema based on a foundation laid by George Palade's classical electron microscopic characterization of the cellular secretory pathway. Secretory proteins are synthesized and folded in the endoplasmic reticulum (ER). They then gather at specialized subdomains of the ER called ER exit sites, marked by coat protein complex II (COPII) proteins. In the conventional model, proteins are packaged into COPII-coated vesicles, which detach from the ER and fuse with the next compartment, the ERGIC (ER-Golgi intermediate compartment) and thence on through the Golgi apparatus to their final destination. Materials that need to be retrieved are packaged into a similar class of vesicles called COPI-coated vesicles for their retrograde transport to the ER.

Coat protein complex II (COPII)-coated vesicle biogenesis. Cytosolic machinery assembles in a hierarchical manner at an endoplasmic reticulum (ER) exit site (ERES). The small GTPase Sar1 is recruited to the ER membrane. This in turn recruits an inner COPII coat of Sec23/Sec24. Secretory cargo in the ER binds to cargo receptors, which bind to Sec24. The Sar1-Sec23/24 “prebudding” complex recruits an outer COPII coat of Sec13/31 that sculpts the site into a vesicle. The vesicle buds off of the ER and forms a 60-nm coated vesicle. Vesicle uncoating exposes machinery required for vesicle targeting and fusion to a downstream compartment.
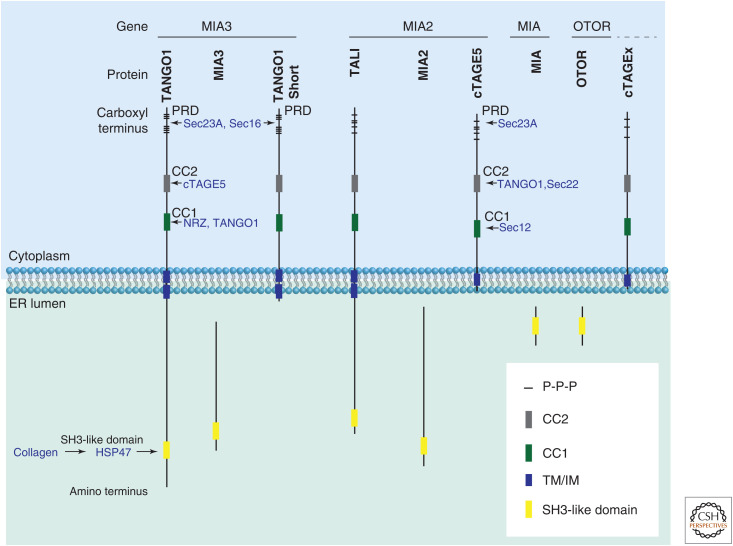
The transport and Golgi organization 1 (TANGO1) family of proteins. Mammalian TANGO1 family of proteins. Full-length TANGO1 or TANGO1-like (TALI) consists of an endoplasmic reticulum (ER)-lumenal part, with extensive regions that are predicted to be intrinsically disordered and an SH3-like domain. The cytoplasmic face of the ER membrane contains coiled coils (CC1 and CC2) and a carboxy-terminal proline-rich domain (PRD). Secreted isoforms MIA and otoraplin are almost exclusively an SH3-like domain. Other paralogs and their isoforms include TANGO1-Short, cTAGE5, and several predicted cTAGEx proteins (where x is 2, 4, 6, 8, 9, and 15). Each domain interacts with a specific set of proteins as indicated.

TANGO1 (transport and Golgi organization 1) stabilizes a transient tunnel between the endoplasmic reticulum (ER) and the ER-Golgi intermediate compartment (ERGIC). TANGO1 at the base of nascent bud, binds to multiple Sec23/24 and promotes a cylindrical coat protein complex II (COPII) assembly. Via the NRZ tether, TANGO1 holds the ERGIC (pink membranes) in place at the ER exit sites (ERES). This could result in a transient continuity generated between the ER (blue) and the ERGIC (pink).
Similar articles
-
COPII and the regulation of protein sorting in mammals.Nat Cell Biol. 2011 Dec 22;14(1):20-8. doi: 10.1038/ncb2390. Nat Cell Biol. 2011. PMID: 22193160 Review.
-
Protein export at the ER: loading big collagens into COPII carriers.EMBO J. 2011 Aug 31;30(17):3475-80. doi: 10.1038/emboj.2011.255. EMBO J. 2011. PMID: 21878990 Free PMC article. Review.
-
Vesicle-mediated export from the ER: COPII coat function and regulation.Biochim Biophys Acta. 2013 Nov;1833(11):2464-72. doi: 10.1016/j.bbamcr.2013.02.003. Epub 2013 Feb 15. Biochim Biophys Acta. 2013. PMID: 23419775 Free PMC article. Review.
-
Tango1 coordinates the formation of endoplasmic reticulum/Golgi docking sites to mediate secretory granule formation.J Biol Chem. 2019 Dec 20;294(51):19498-19510. doi: 10.1074/jbc.RA119.011063. Epub 2019 Nov 5. J Biol Chem. 2019. PMID: 31690624 Free PMC article.
-
TANGO1 assembles into rings around COPII coats at ER exit sites.J Cell Biol. 2017 Apr 3;216(4):901-909. doi: 10.1083/jcb.201608080. Epub 2017 Mar 9. J Cell Biol. 2017. PMID: 28280121 Free PMC article.
Cited by
-
Adaptations of membrane trafficking in cancer and tumorigenesis.J Cell Sci. 2024 May 15;137(10):jcs260943. doi: 10.1242/jcs.260943. Epub 2024 May 21. J Cell Sci. 2024. PMID: 38770683 Free PMC article. Review.
-
TFG regulates inner COPII coat recruitment to facilitate anterograde secretory protein transport.Mol Biol Cell. 2024 Aug 1;35(8):ar113. doi: 10.1091/mbc.E24-06-0282. Epub 2024 Jul 10. Mol Biol Cell. 2024. PMID: 38985515 Free PMC article.
-
Editorial: Homeostatic regulation of protein synthesis, folding and secretion by stress response pathways in eukaryotes.Front Cell Dev Biol. 2023 Sep 13;11:1282272. doi: 10.3389/fcell.2023.1282272. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37779897 Free PMC article. No abstract available.
-
The secretory pathway in Tetrahymena is organized for efficient constitutive secretion at ciliary pockets.iScience. 2024 Oct 9;27(11):111123. doi: 10.1016/j.isci.2024.111123. eCollection 2024 Nov 15. iScience. 2024. PMID: 39498308 Free PMC article.
-
Targets and Potential Mechanism of Chondroitin Sulfate A-selenium Nanoparticle on Kashin-Beck Disease Chondrocytes.Biol Trace Elem Res. 2025 Mar 26. doi: 10.1007/s12011-025-04584-3. Online ahead of print. Biol Trace Elem Res. 2025. PMID: 40138104
References
-
- Amodio G, Renna M, Paladino S, Venturi C, Tacchetti C, Moltedo O, Franceschelli S, Mallardo M, Bonatti S, Remondelli P. 2009. Endoplasmic reticulum stress reduces the export from the ER and alters the architecture of post-ER compartments. Int J Biochem Cell Biol 41: 2511–2521. 10.1016/j.biocel.2009.08.006 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases