

Chromatin compaction precedes apoptosis in developing neurons
- PMID: 35941180
- PMCID: PMC9359995
- DOI: 10.1038/s42003-022-03704-2
Chromatin compaction precedes apoptosis in developing neurons
Abstract
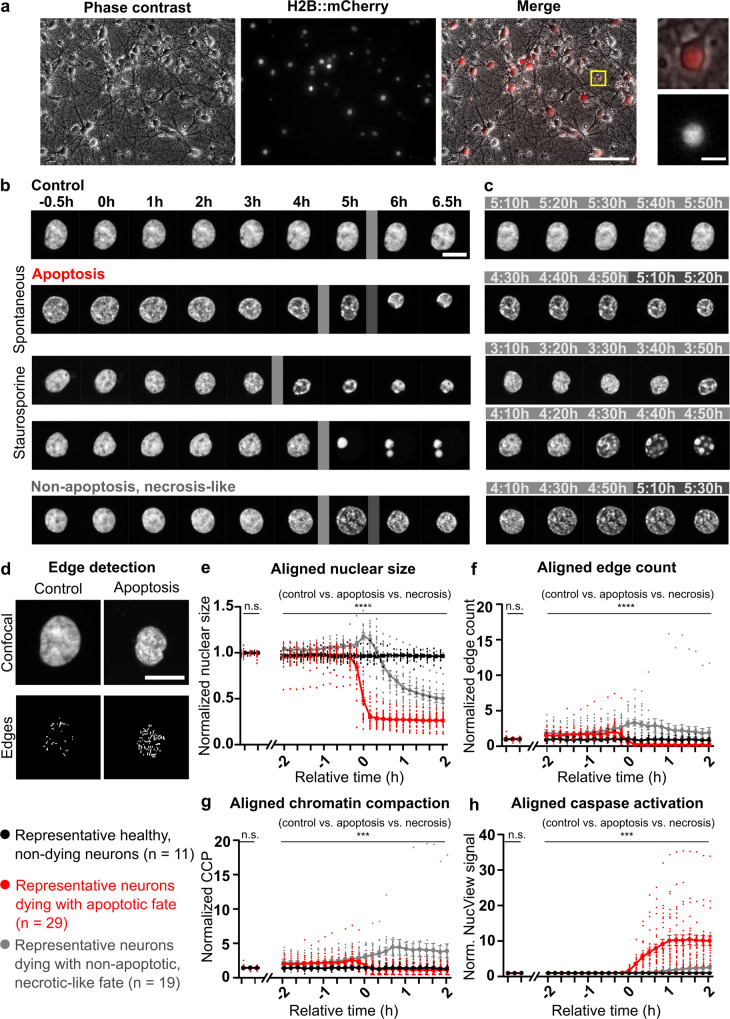
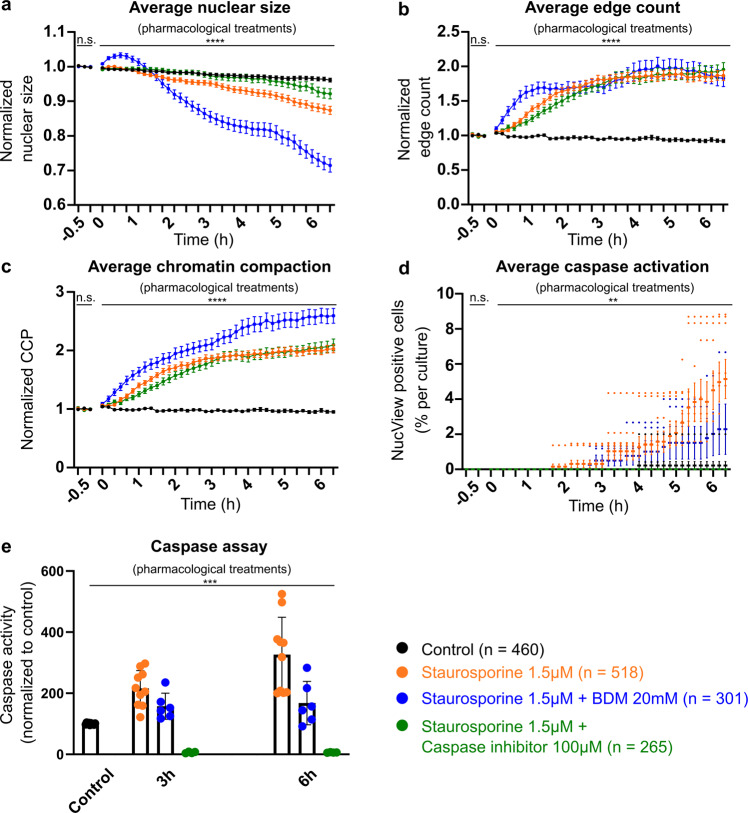
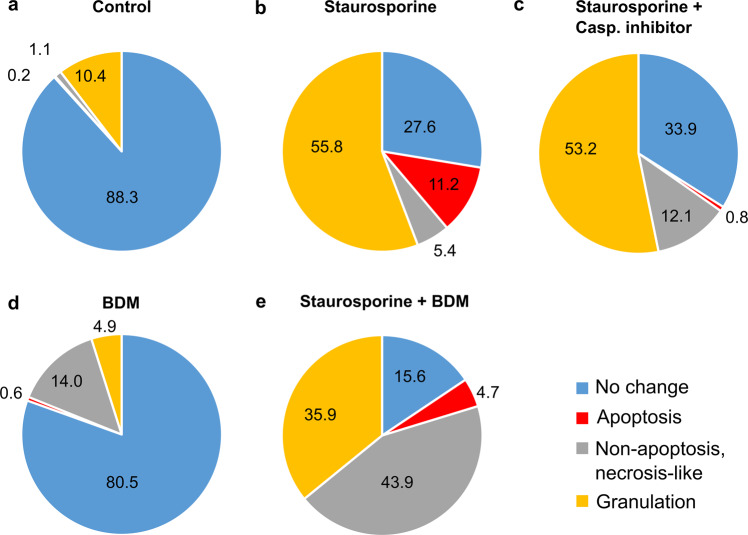
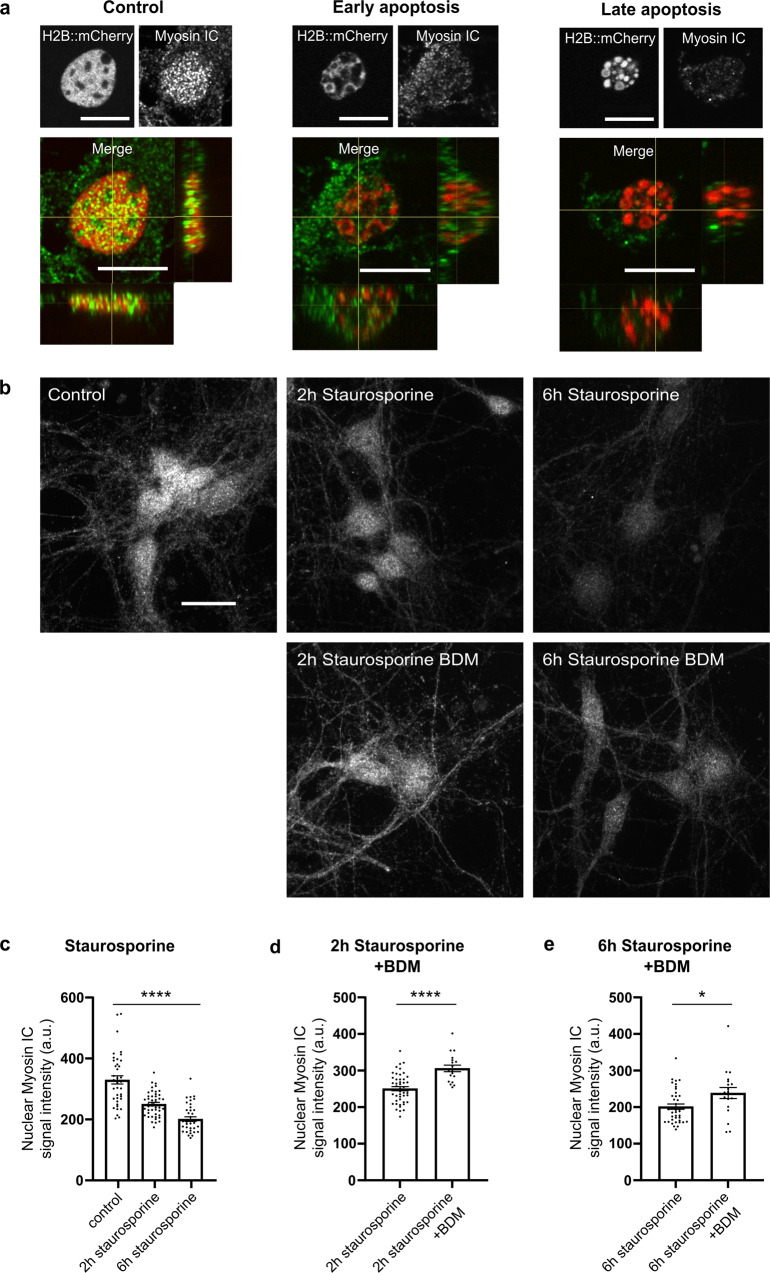
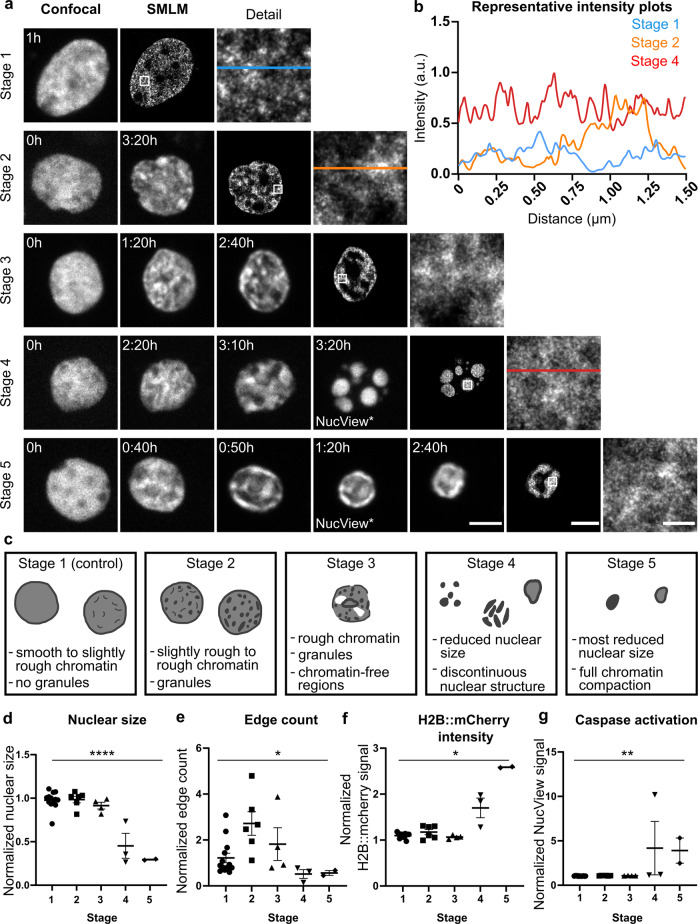
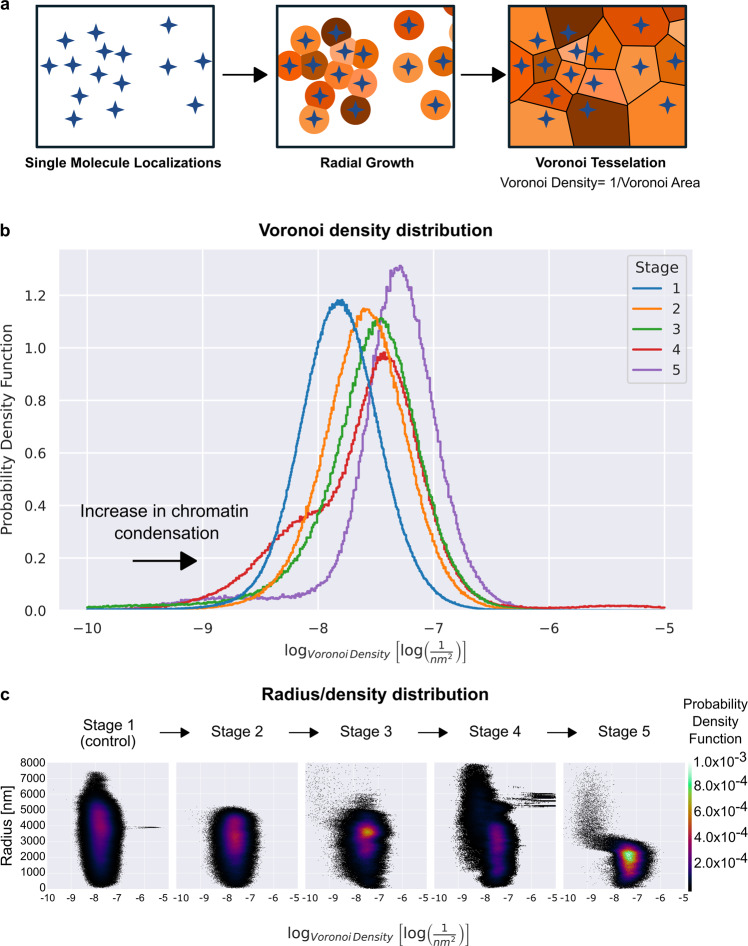
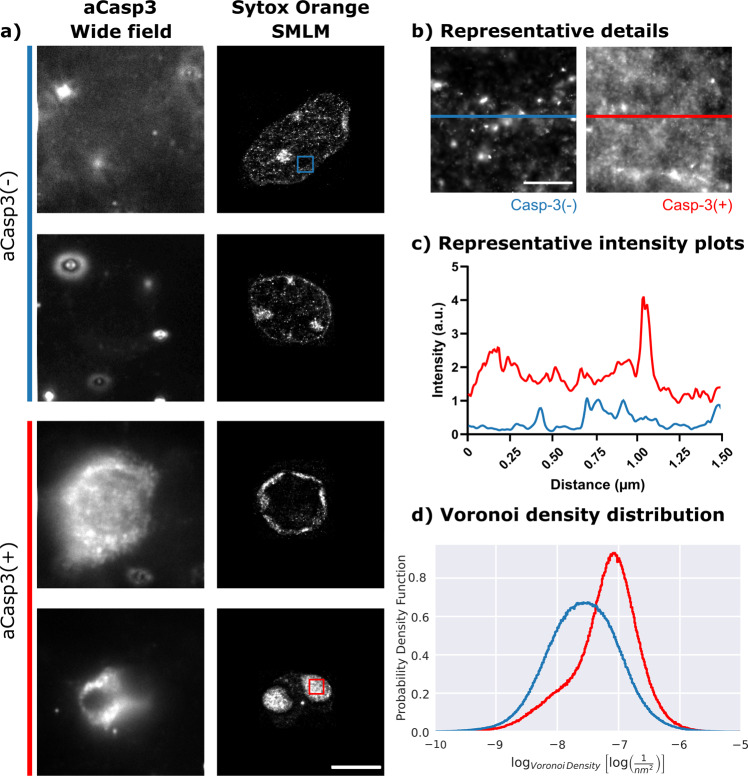
While major changes in cellular morphology during apoptosis have been well described, the subcellular changes in nuclear architecture involved in this process remain poorly understood. Imaging of nucleosomes in cortical neurons in vitro before and during apoptosis revealed that chromatin compaction precedes the activation of caspase-3 and nucleus shrinkage. While this early chromatin compaction remained unaffected by pharmacological blockade of the final execution of apoptosis through caspase-3 inhibition, interfering with the chromatin dynamics by modulation of actomyosin activity prevented apoptosis, but resulted in necrotic-like cell death instead. With super-resolution imaging at different phases of apoptosis in vitro and in vivo, we demonstrate that chromatin compaction occurs progressively and can be classified into five stages. In conclusion, we show that compaction of chromatin in the neuronal nucleus precedes apoptosis execution. These early changes in chromatin structure critically affect apoptotic cell death and are not part of the final execution of the apoptotic process in developing cortical neurons.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
