

CCR9 axis inhibition enhances hepatic migration of plasmacytoid DCs and protects against liver injury
- PMID: 35943802
- PMCID: PMC9536268
- DOI: 10.1172/jci.insight.159910
CCR9 axis inhibition enhances hepatic migration of plasmacytoid DCs and protects against liver injury
Abstract
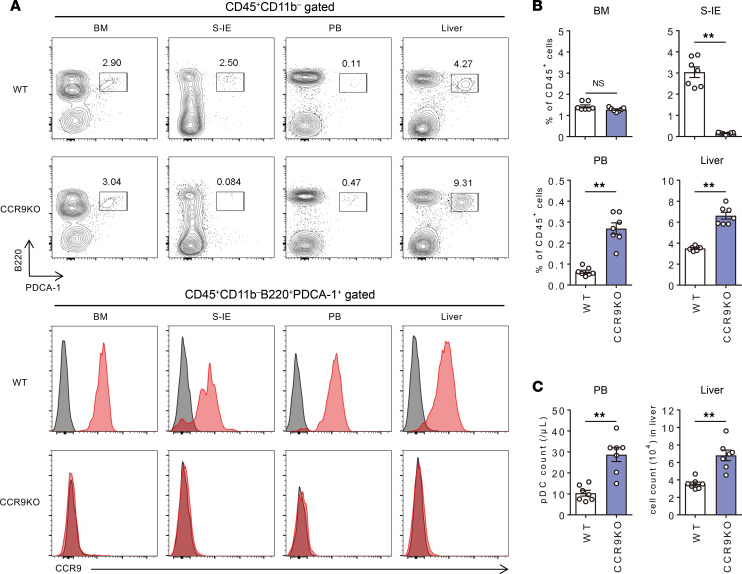
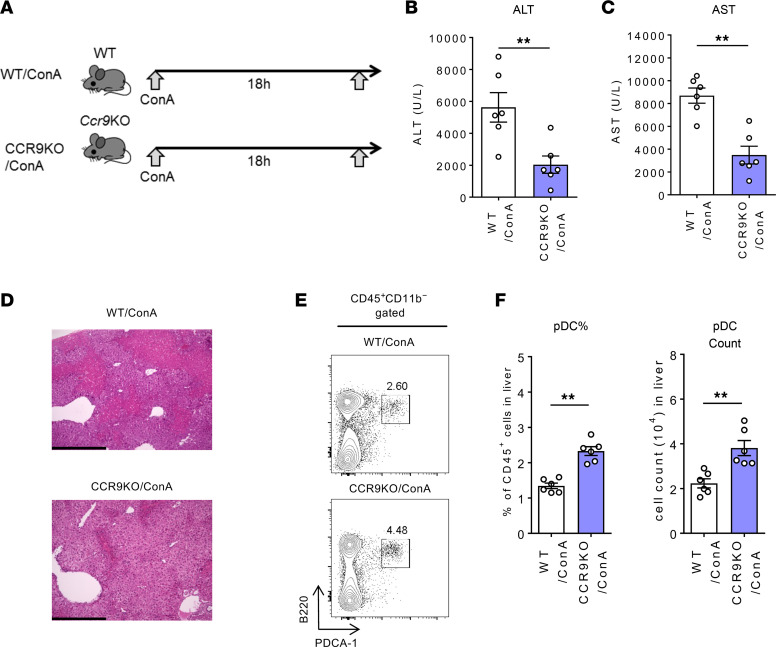
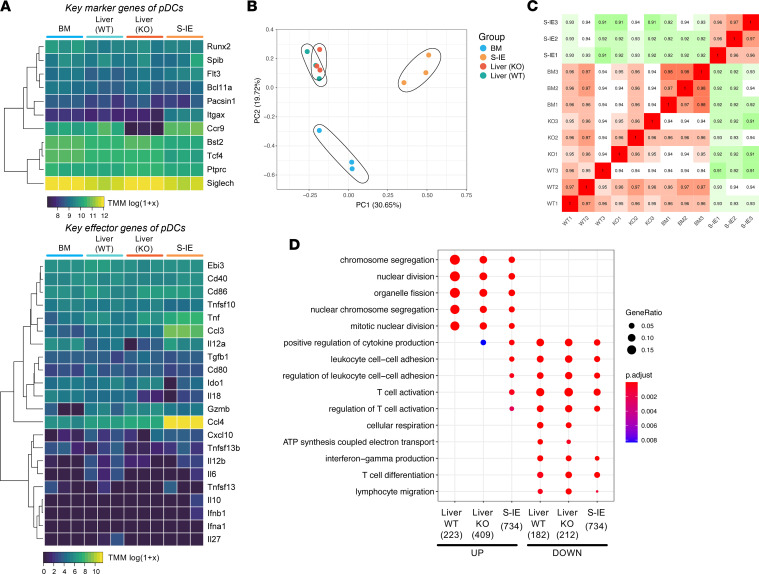
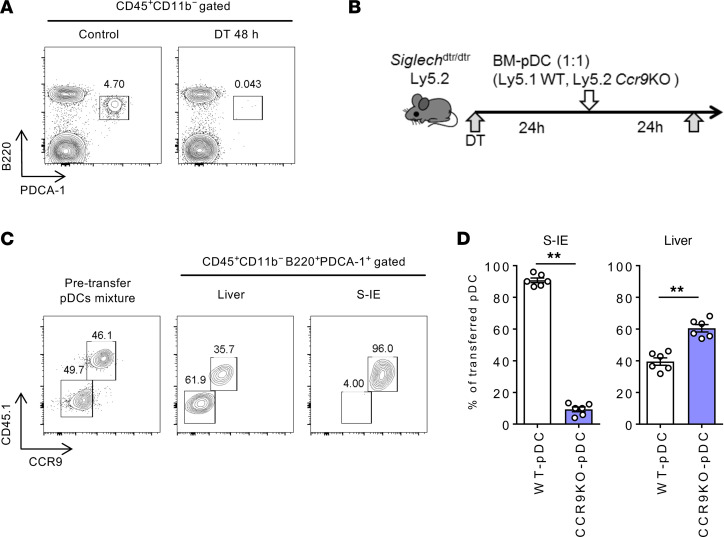
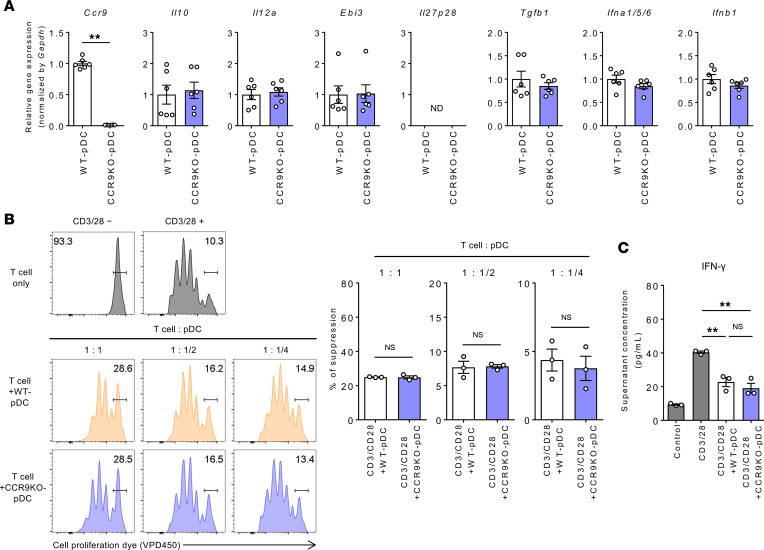
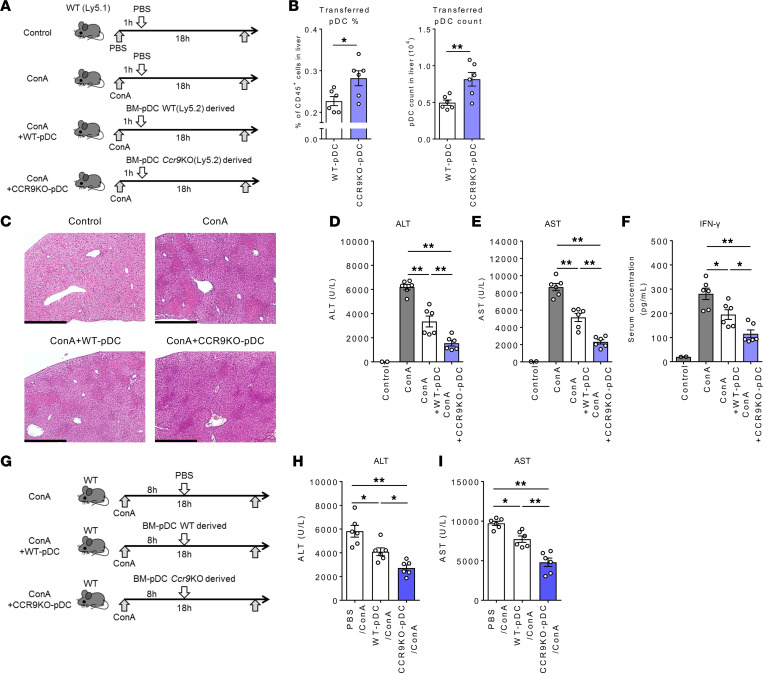
Plasmacytoid dendritic cells (pDCs) perform dual proinflammatory and immunosuppressive roles. We recently reported the potential of pDC therapy for treatment of intractable acute liver failure. However, establishment of efficient methods to deliver pDCs to the liver is essential for future clinical therapeutic applications. The present study demonstrates a higher abundance of liver and peripheral blood pDCs in mice lacking C-C motif chemokine receptor 9 (CCR9), a pDC gut-homing receptor, than in WT mice. Adoptive transfer of Ccr9-/- pDCs resulted in a higher efficiency of migration to the liver than WT pDCs did, while WT pDCs migrated efficiently to the original target organ, the small intestine. Further, Ccr9-/- pDCs consistently migrated efficiently to livers with concanavalin A-induced inflammation, and exerted a more effective immunosuppressive effect, resulting in better protection against acute liver inflammation than that demonstrated by WT pDCs. These findings highlight the therapeutic potential of the manipulation of the CCR9 axis as an approach to improve migration of immunosuppressive pDCs to the liver in order to exploit their beneficial effects in acute liver disease.
Keywords: Chemokines; Dendritic cells; Hepatitis; Hepatology; Immunology.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
