

N-Terminal Acetylation of α-Synuclein Slows down Its Aggregation Process and Alters the Morphology of the Resulting Aggregates
- PMID: 35944093
- PMCID: PMC9454101
- DOI: 10.1021/acs.biochem.2c00104
N-Terminal Acetylation of α-Synuclein Slows down Its Aggregation Process and Alters the Morphology of the Resulting Aggregates
Abstract
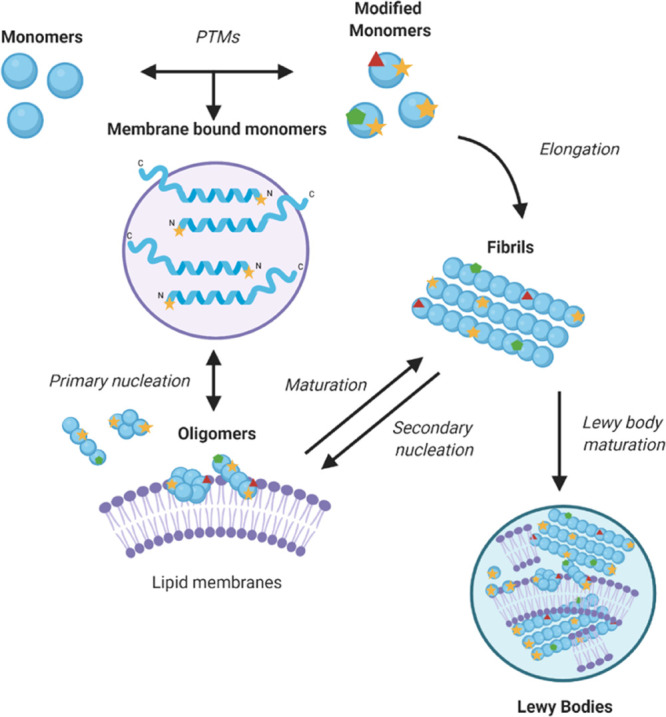
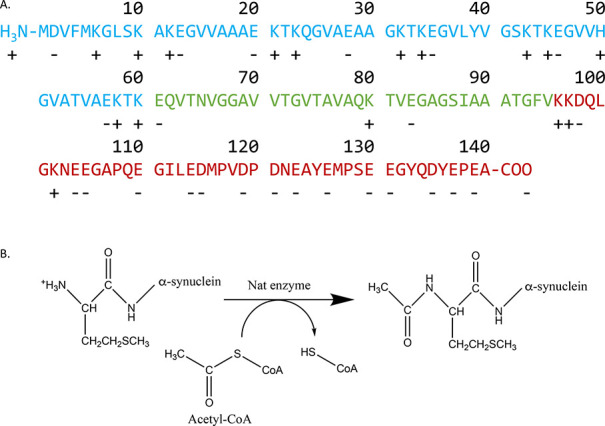
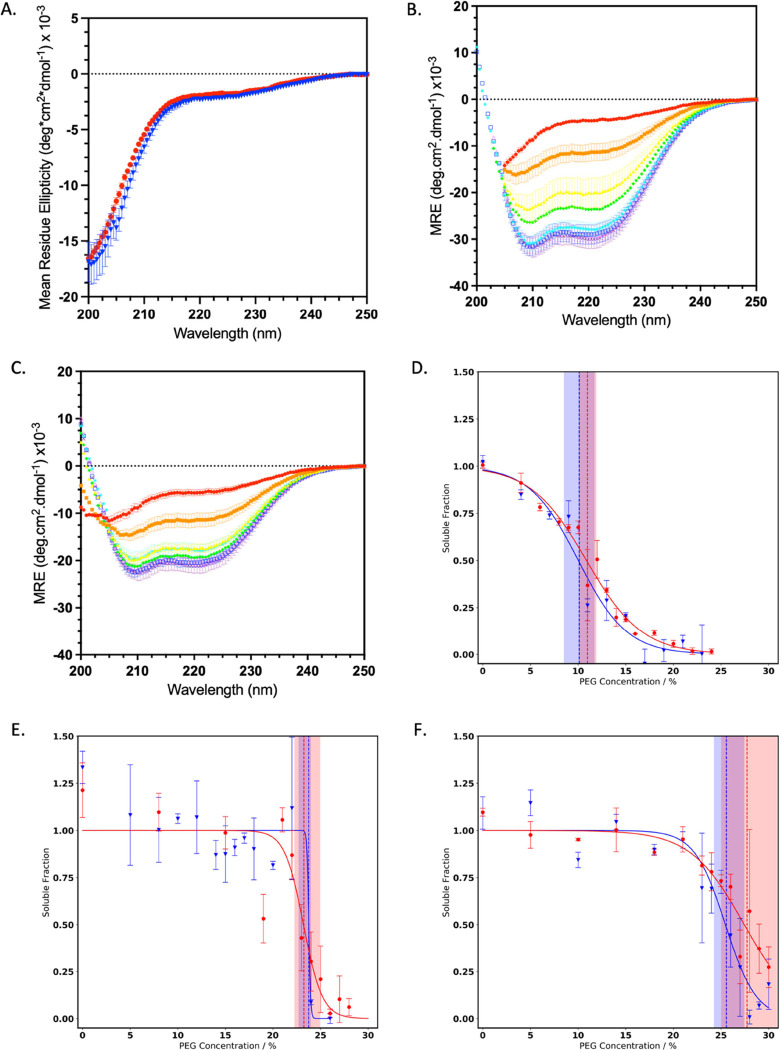
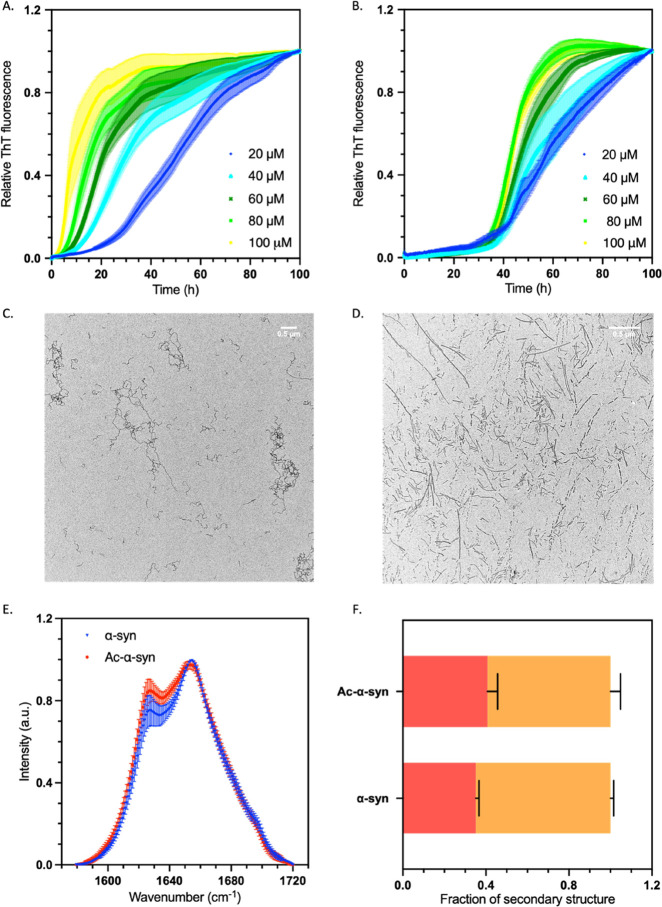
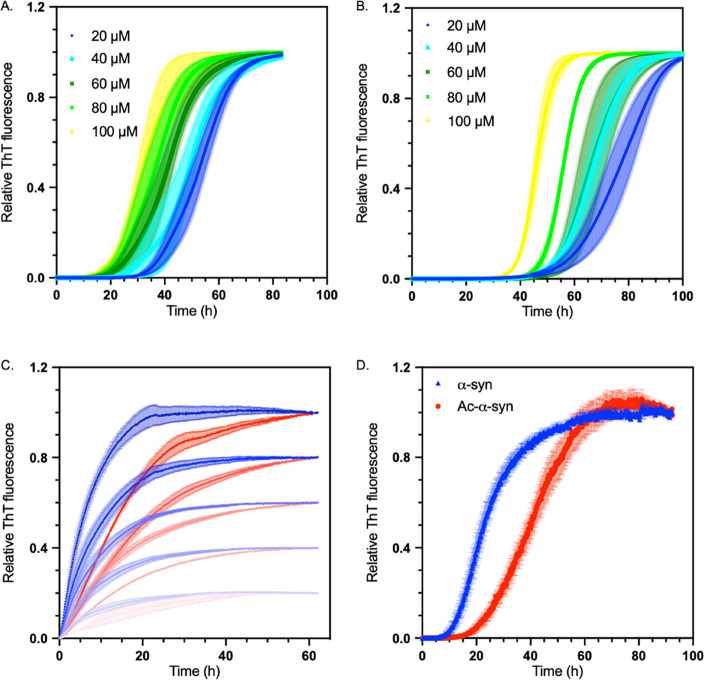
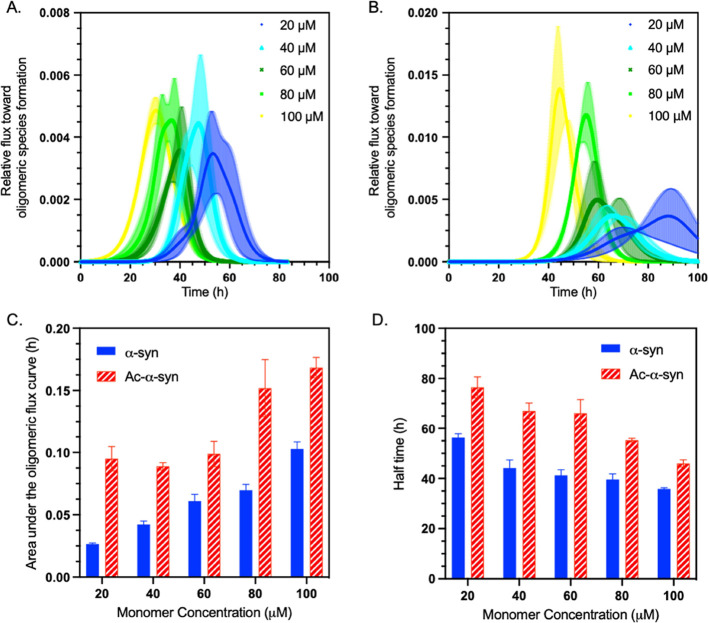
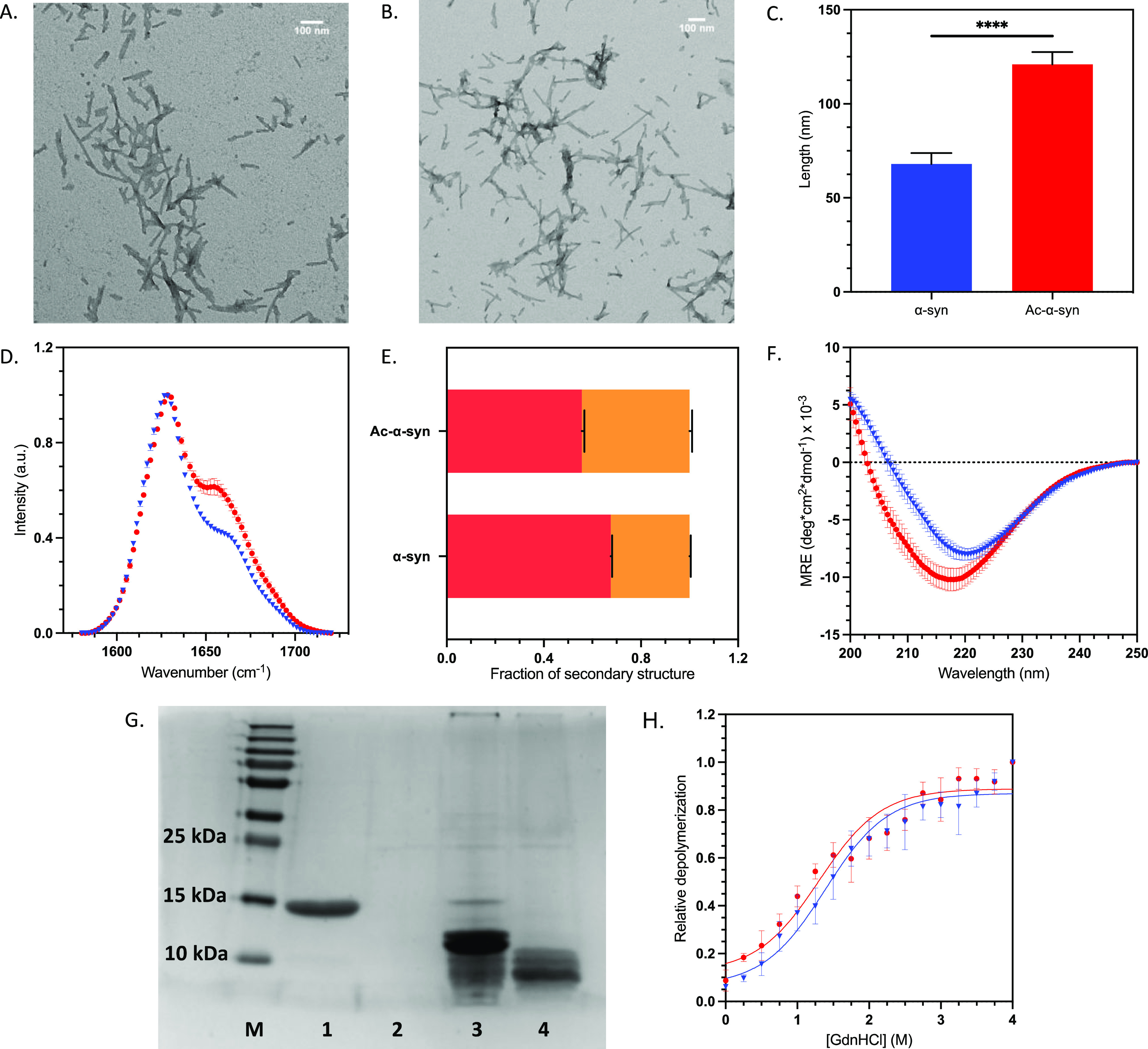
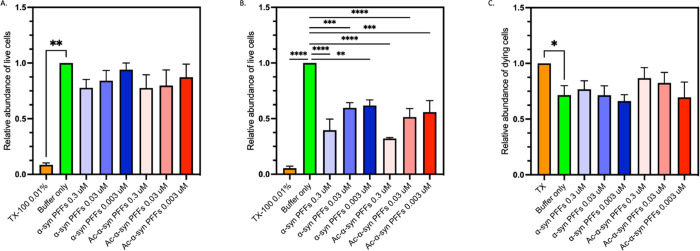
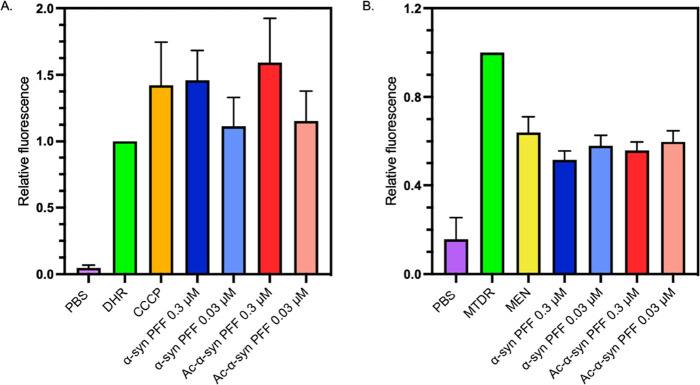
Parkinson's disease is associated with the aberrant aggregation of α-synuclein. Although the causes of this process are still unclear, post-translational modifications of α-synuclein are likely to play a modulatory role. Since α-synuclein is constitutively N-terminally acetylated, we investigated how this post-translational modification alters the aggregation behavior of this protein. By applying a three-pronged aggregation kinetics approach, we observed that N-terminal acetylation results in a reduced rate of lipid-induced aggregation and slows down both elongation and fibril-catalyzed aggregate proliferation. An analysis of the amyloid fibrils produced by the aggregation process revealed different morphologies for the acetylated and non-acetylated forms in both lipid-induced aggregation and seed-induced aggregation assays. In addition, we found that fibrils formed by acetylated α-synuclein exhibit a lower β-sheet content. These findings indicate that N-terminal acetylation of α-synuclein alters its lipid-dependent aggregation behavior, reduces its rate of in vitro aggregation, and affects the structural properties of its fibrillar aggregates.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Ray Dorsey E.; Elbaz A.; Nichols E.; Abd-Allah F.; Abdelalim A.; Adsuar J. C.; Ansha M. G.; Brayne C.; Choi J. Y. J.; Collado-Mateo D.; Dahodwala N.; Do H. P.; Edessa D.; Endres M.; Fereshtehnejad S. M.; Foreman K. J.; Gankpe F. G.; Gupta R.; Hankey G. J.; Hay S. I.; Hegazy M. I.; Hibstu D. T.; Kasaeian A.; Khader Y.; Khalil I.; Khang Y. H.; Kim Y. J.; Kokubo Y.; Logroscino G.; Massano J.; Ibrahim N. M.; Mohammed M. A.; Mohammadi A.; Moradi-Lakeh M.; Naghavi M.; Nguyen B. T.; Nirayo Y. L.; Ogbo F. A.; Owolabi M. O.; Pereira D. M.; Postma M. J.; Qorbani M.; Rahman M. A.; Roba K. T.; Safari H.; Safiri S.; Satpathy M.; Sawhney M.; Shafieesabet A.; Shiferaw M. S.; Smith M.; Szoeke C. E. I.; Tabarés-Seisdedos R.; Truong N. T.; Ukwaja K. N.; Venketasubramanian N.; Villafaina S.; Weldegwergs K. G.; Westerman R.; Wijeratne T.; Winkler A. S.; Xuan B. T.; Yonemoto N.; Feigin V. L.; Vos T.; Murray C. J. L. Global, regional, and national burden of Parkinson’s disease, 1990–2016: a systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2018, 17, 939–953. 10.1016/S1474-4422(18)30295-3. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
