

Covalent Proteomimetic Inhibitor of the Bacterial FtsQB Divisome Complex
- PMID: 35945166
- PMCID: PMC9413201
- DOI: 10.1021/jacs.2c06304
Covalent Proteomimetic Inhibitor of the Bacterial FtsQB Divisome Complex
Abstract
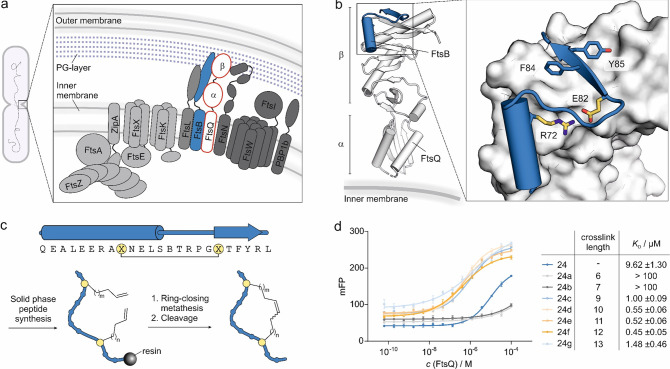
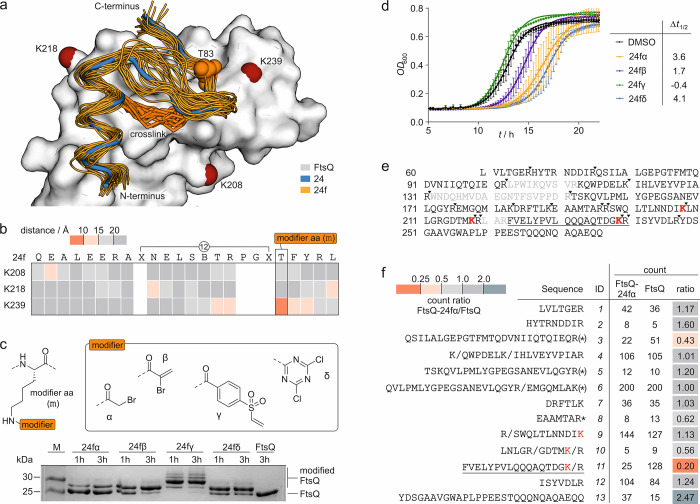
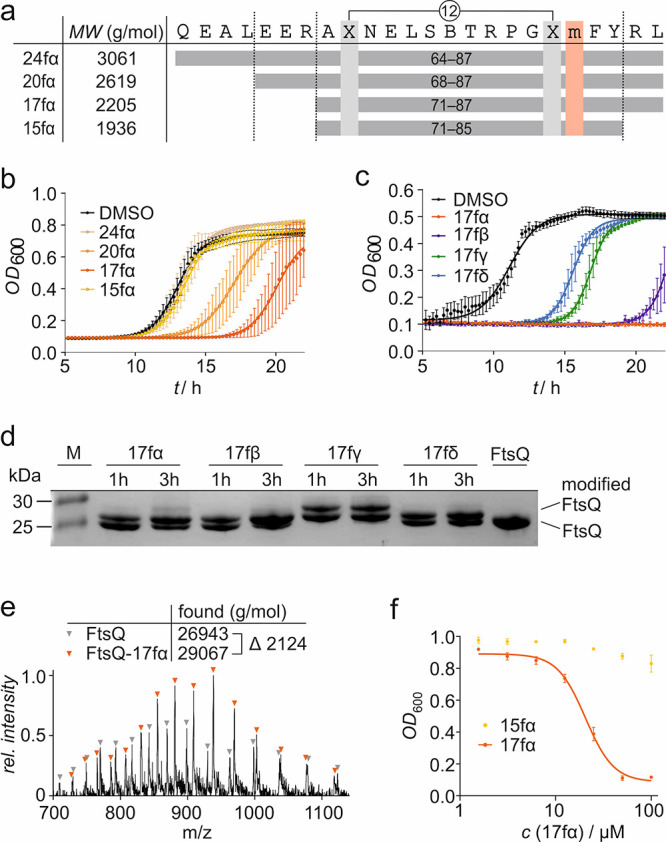
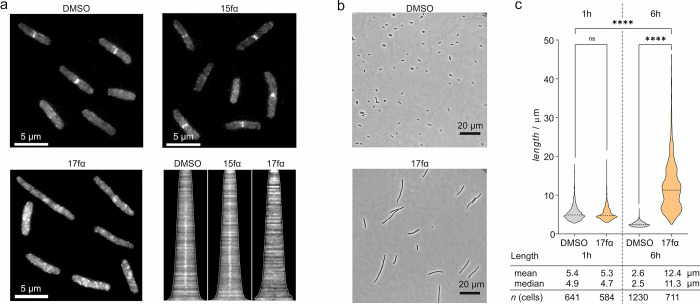
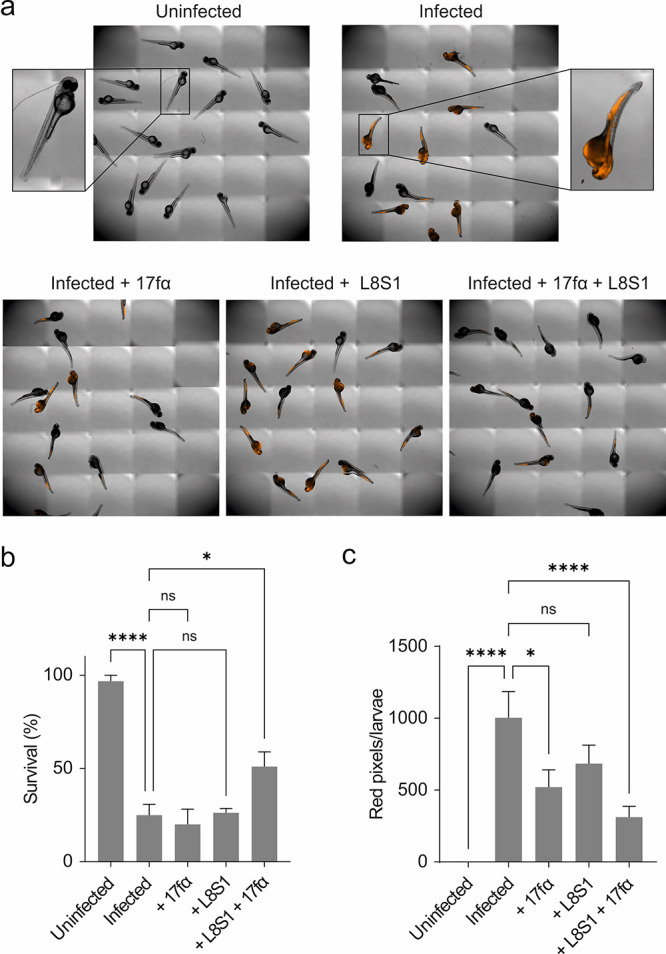
The use of antibiotics is threatened by the emergence and spread of multidrug-resistant strains of bacteria. Thus, there is a need to develop antibiotics that address new targets. In this respect, the bacterial divisome, a multi-protein complex central to cell division, represents a potentially attractive target. Of particular interest is the FtsQB subcomplex that plays a decisive role in divisome assembly and peptidoglycan biogenesis in E. coli. Here, we report the structure-based design of a macrocyclic covalent inhibitor derived from a periplasmic region of FtsB that mediates its binding to FtsQ. The bioactive conformation of this motif was stabilized by a customized cross-link resulting in a tertiary structure mimetic with increased affinity for FtsQ. To increase activity, a covalent handle was incorporated, providing an inhibitor that impedes the interaction between FtsQ and FtsB irreversibly. The covalent inhibitor reduced the growth of an outer membrane-permeable E. coli strain, concurrent with the expected loss of FtsB localization, and also affected the infection of zebrafish larvae by a clinical E. coli strain. This first-in-class inhibitor of a divisome protein-protein interaction highlights the potential of proteomimetic molecules as inhibitors of challenging targets. In particular, the covalent mode-of-action can serve as an inspiration for future antibiotics that target protein-protein interactions.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
